

Mycobacterium tuberculosis-specific CD8+ T cells are functionally and phenotypically different between latent infection and active disease
- PMID: 23456989
- PMCID: PMC6535091
- DOI: 10.1002/eji.201243262
Mycobacterium tuberculosis-specific CD8+ T cells are functionally and phenotypically different between latent infection and active disease
Abstract
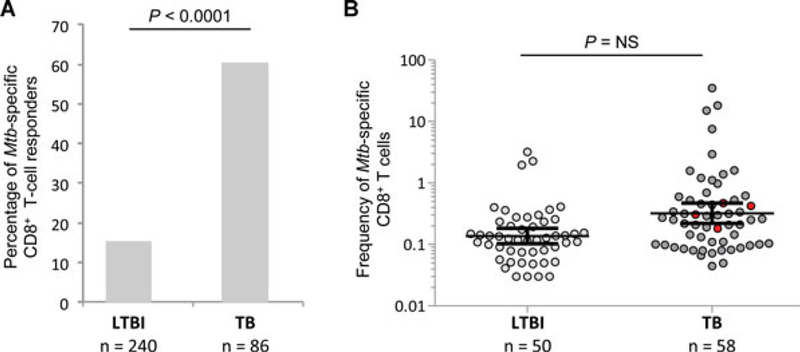
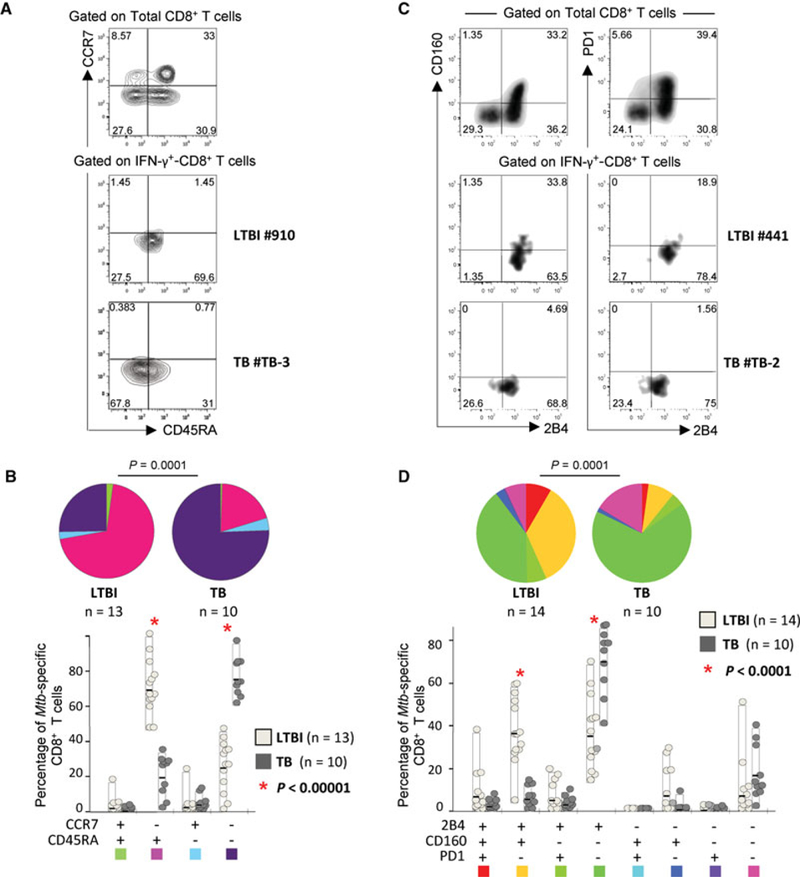
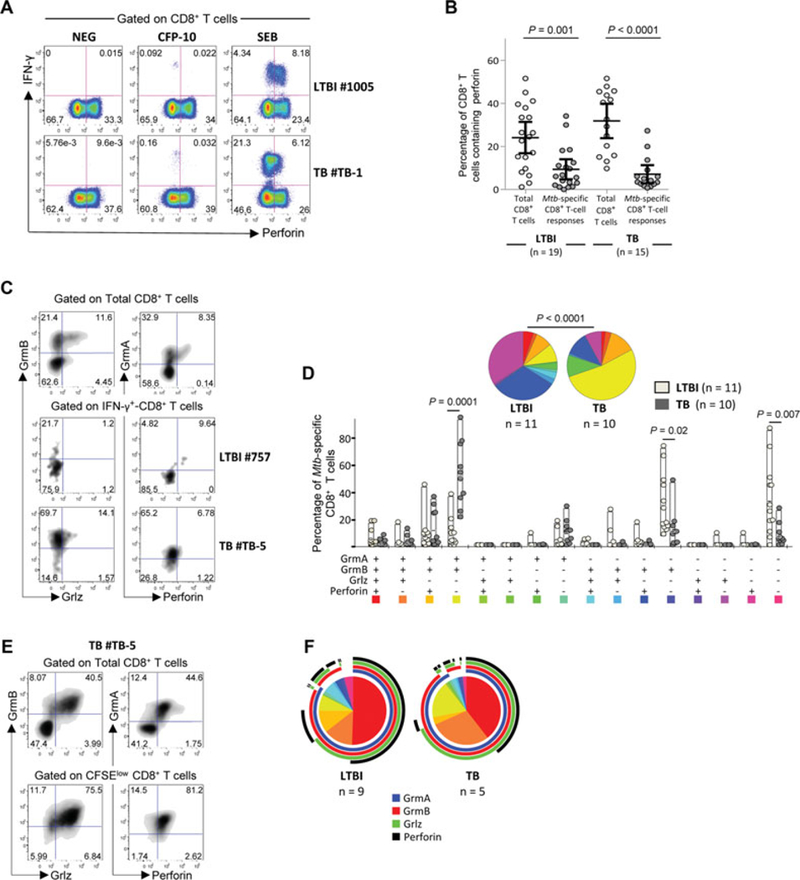
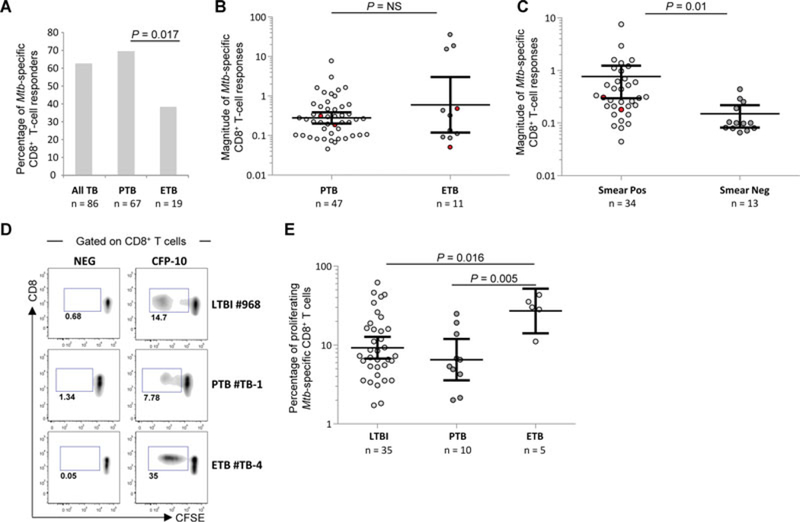
Protective immunity to Mycobacterium tuberculosis (Mtb) remains poorly understood and the role of Mtb-specific CD8(+) T cells is controversial. Here we performed a broad phenotypic and functional characterization of Mtb-specific CD8(+) T cells in 326 subjects with latent Mtb infection (LTBI) or active TB disease (TB). Mtb-specific CD8(+) T cells were detected in most (60%) TB patients and few (15%) LTBI subjects but were of similar magnitude. Mtb-specific CD8(+) T cells in LTBI subjects were mostly T EMRA cells (CD45RA(+) CCR7(-)), coexpressing 2B4 and CD160, and in TB patients were mostly TEM cells (CD45RA(-) CCR7(-)), expressing 2B4 but lacking PD-1 and CD160. The cytokine profile was not significantly different in both groups. Furthermore, Mtb-specific CD8(+) T cells expressed low levels of perforin and granulysin but contained granzymes A and B. However, in vitro-expanded Mtb-specific CD8(+) T cells expressed perforin and granulysin. Finally, Mtb-specific CD8(+) T-cell responses were less frequently detected in extrapulmonary TB compared with pulmonary TB patients. Mtb-specific CD8(+) T-cell proliferation was also greater in patients with extrapulmonary compared with pulmonary TB. Thus, the activity of Mtb infection and clinical presentation are associated with distinct profiles of Mtb-specific CD8(+) T-cell responses. These results provide new insights in the interaction between Mtb and the host immune response.
© 2013 WILEY-VCH Verlag GmbH & Co. KGaA, Weinheim.
Conflict of interest statement
Conflict of interest: The authors declare no financial or commercial conflict of interest.
Figures
References
-
- Koul A, Arnoult E, Lounis N, Guillemont J and Andries K, The challenge of new drug discovery for tuberculosis. Nature 2011. 469: 483–490. - PubMed
-
- Winthrop KL, Risk and prevention of tuberculosis and other serious opportunistic infections associated with the inhibition of tumor necrosis factor. Nat. Clin. Pract. Rheumatol 2006. 2: 602–610. - PubMed
-
- Bumbacea D, Arend SM, Eyuboglu F, Fishman JA, Goletti D, Ison MG, Jones CE et al. , The risk of tuberculosis in transplant candidates and recipients: a TBNET consensus statement. Eur. Respir. J 2012. 40: 990–1013. - PubMed
-
- Granich R, Akolo C, Gunneberg C, Getahun H, Williams P and Williams B, Prevention of tuberculosis in people living with HIV. Clin. Infect. Dis 2010. 50(Suppl 3): S215–S222. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Research Materials
