

The major outer sheath protein (Msp) of Treponema denticola has a bipartite domain architecture and exists as periplasmic and outer membrane-spanning conformers
- PMID: 23457251
- PMCID: PMC3624580
- DOI: 10.1128/JB.00078-13
The major outer sheath protein (Msp) of Treponema denticola has a bipartite domain architecture and exists as periplasmic and outer membrane-spanning conformers
Erratum in
- J Bacteriol. 2014 Sep;196(18):3361
Abstract
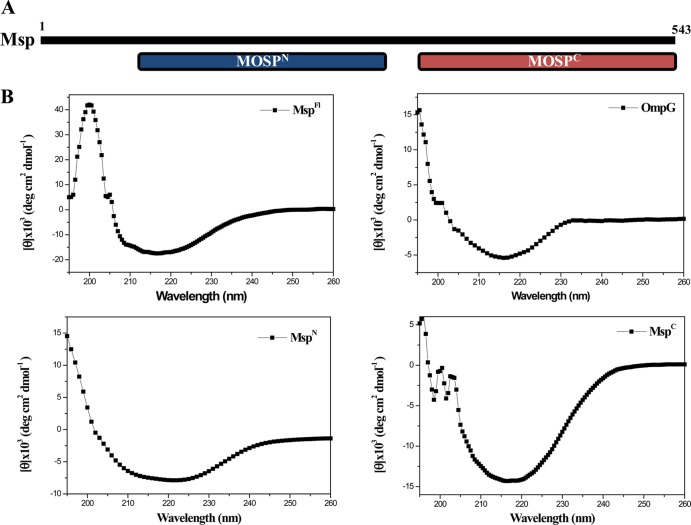
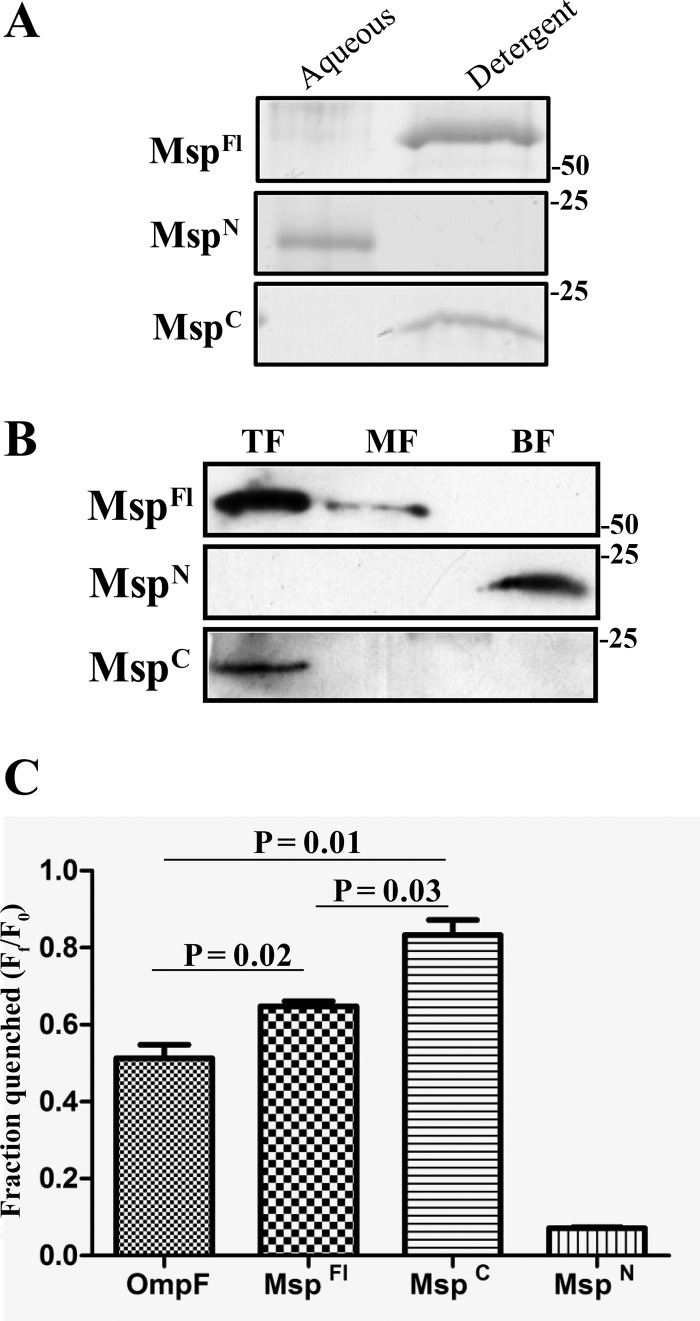
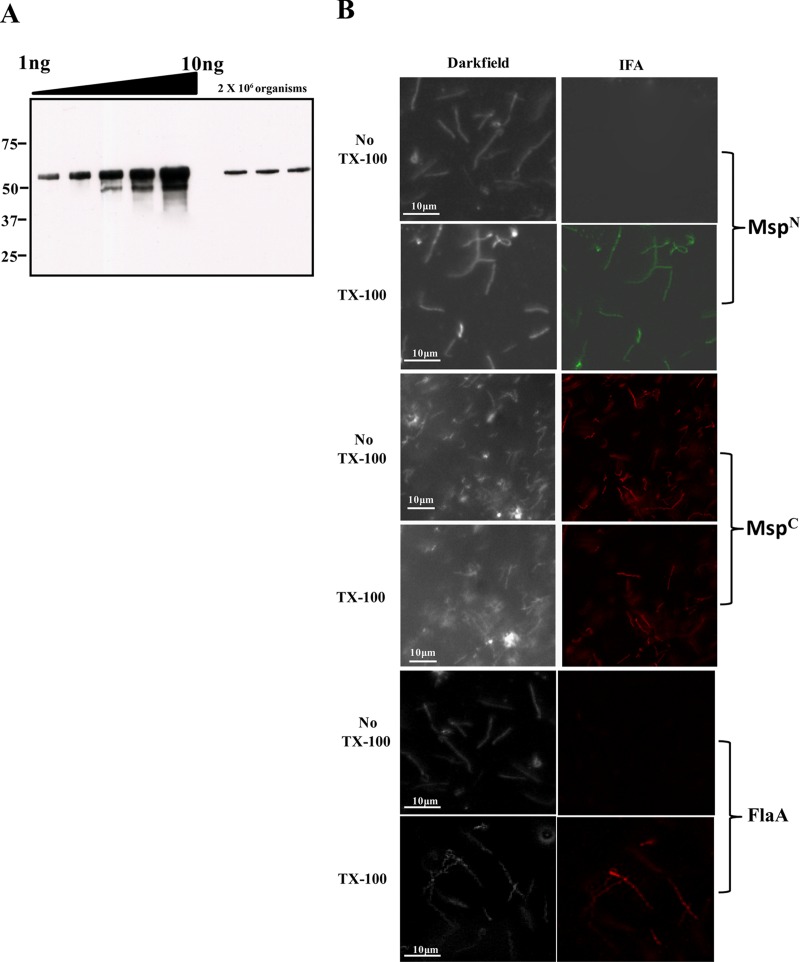
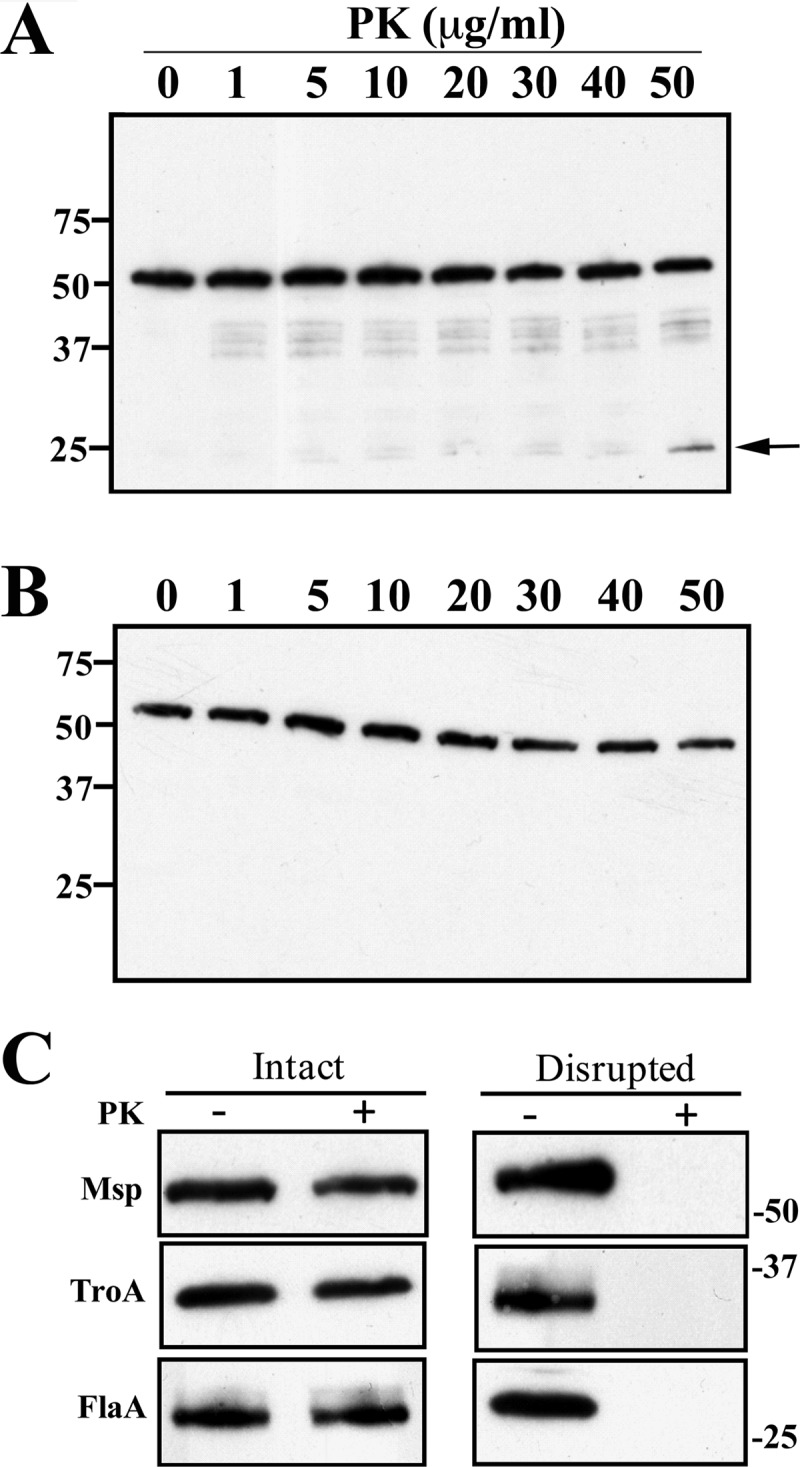
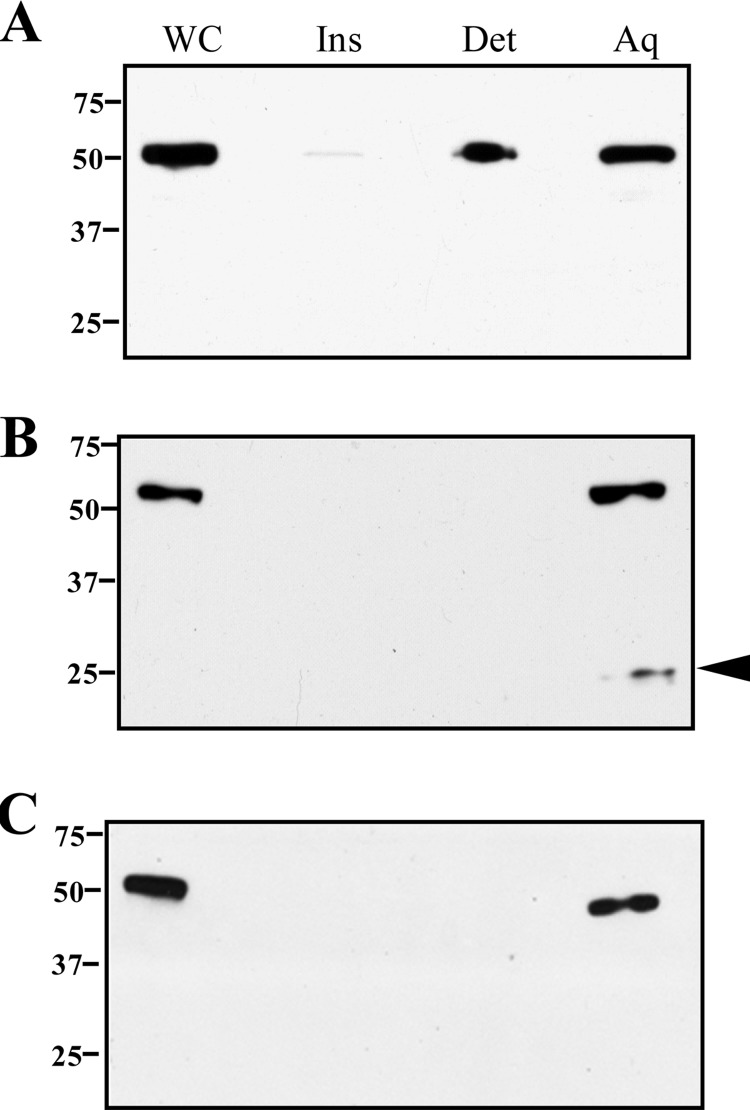
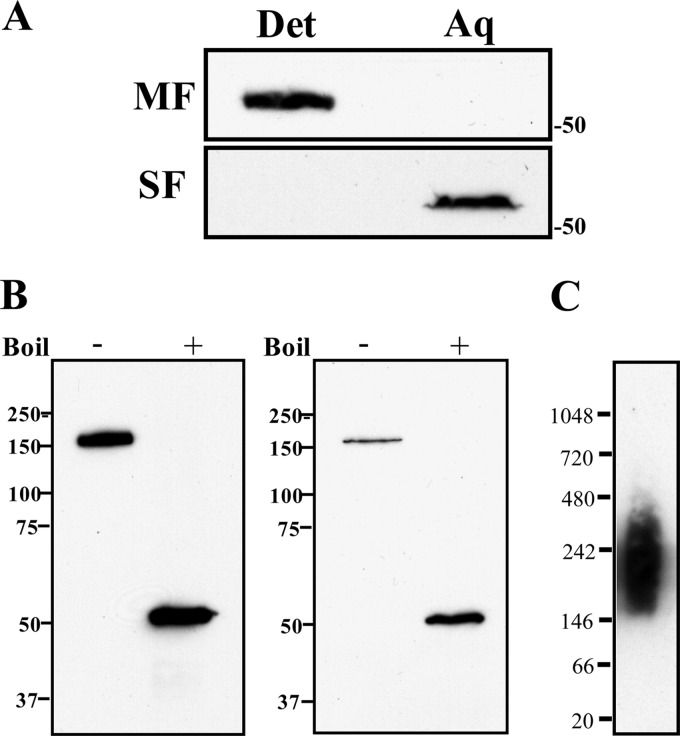
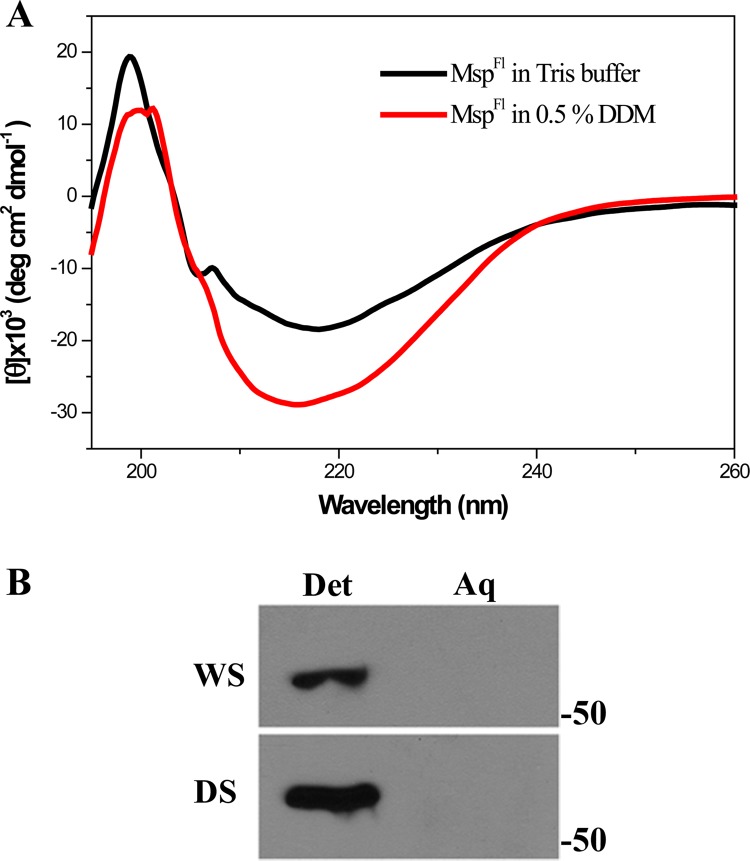
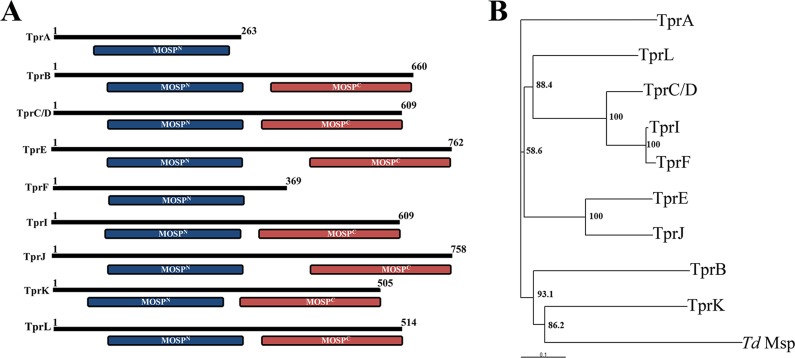
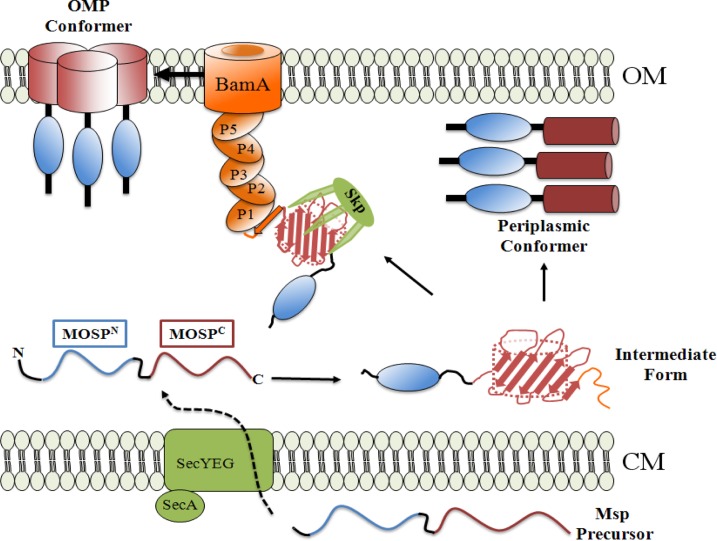
The major outer sheath protein (Msp) is a primary virulence determinant in Treponema denticola, as well as the parental ortholog for the Treponema pallidum repeat (Tpr) family in the syphilis spirochete. The Conserved Domain Database (CDD) server revealed that Msp contains two conserved domains, major outer sheath protein(N) (MOSP(N)) and MOSP(C), spanning residues 77 to 286 and 332 to 543, respectively, within the N- and C-terminal regions of the protein. Circular dichroism (CD) spectroscopy, Triton X-114 (TX-114) phase partitioning, and liposome incorporation demonstrated that full-length, recombinant Msp (Msp(Fl)) and a recombinant protein containing MOSP(C), but not MOSP(N), form amphiphilic, β-sheet-rich structures with channel-forming activity. Immunofluorescence analysis of intact T. denticola revealed that only MOSP(C) contains surface-exposed epitopes. Data obtained using proteinase K accessibility, TX-114 phase partitioning, and cell fractionation revealed that Msp exists as distinct OM-integrated and periplasmic trimers. Msp(Fl) folded in Tris buffer contained slightly less β-sheet structure than detergent-folded Msp(Fl); both forms, however, partitioned into the TX-114 detergent-enriched phase. CDD analysis of the nine Tpr paralogs predicted to be outer membrane proteins (OMPs) revealed that seven have an Msp-like bipartite structure; phylogenetic analysis revealed that the MOSP(N) and MOSP(C) domains of Msp are most closely related to those of TprK. Based upon our collective results, we propose a model whereby a newly exported, partially folded intermediate can be either processed for OM insertion by the β-barrel assembly machinery (BAM) or remain periplasmic, ultimately forming a stable, water-soluble trimer. Extrapolated to T. pallidum, our model enables us to explain how individual Tprs can localize to either the periplasmic (e.g., TprK) or OM (e.g., TprC) compartments.
Figures
References
-
- Holt SC, Ebersole JL. 2006. The oral spirochetes: their ecology and role in the pathogenesis of periodontal disease, p 323–356 In Radolf JD, Lukehart SA. (ed), Pathogenic Treponema. Caister Academic Press, Norwich, United Kingdom
-
- Ellen RP. 2006. Virulence determinants of oral treponemes, p 357–386 In Radolf JD, Lukehart SA. (ed), Pathogenic Treponema. Caister Academic Press, Norwich, United Kingdom
-
- Amano A. 2010. Host-parasite interactions in periodontitis: microbial pathogenicity and innate immunity. Periodontol 2000 54:9–14 - PubMed
-
- Hernandez M, Dutzan N, Garcia-Sesnich J, Abusleme L, Dezerega A, Silva N, Gonzalez FE, Vernal R, Sorsa T, Gamonal J. 2011. Host-pathogen interactions in progressive chronic periodontitis. J. Dent. Res. 90:1164–1170 - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
Research Materials
Miscellaneous
