

Structural biology of the writers, readers, and erasers in mono- and poly(ADP-ribose) mediated signaling
- PMID: 23458732
- PMCID: PMC3726583
- DOI: 10.1016/j.mam.2013.02.002
Structural biology of the writers, readers, and erasers in mono- and poly(ADP-ribose) mediated signaling
Abstract
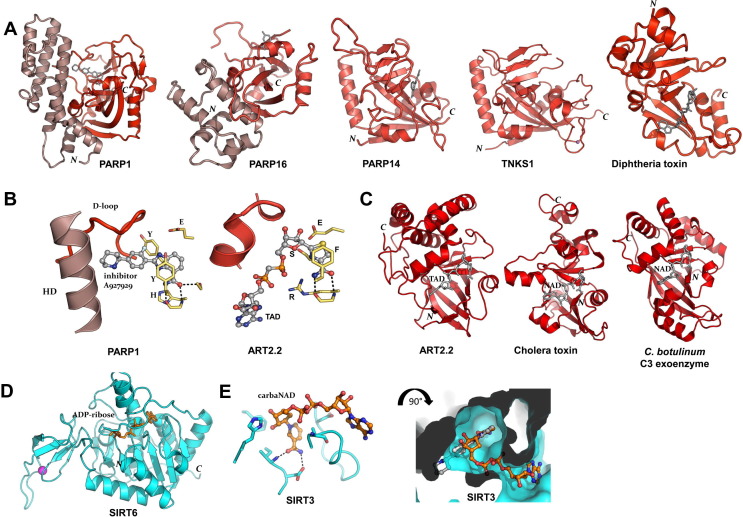
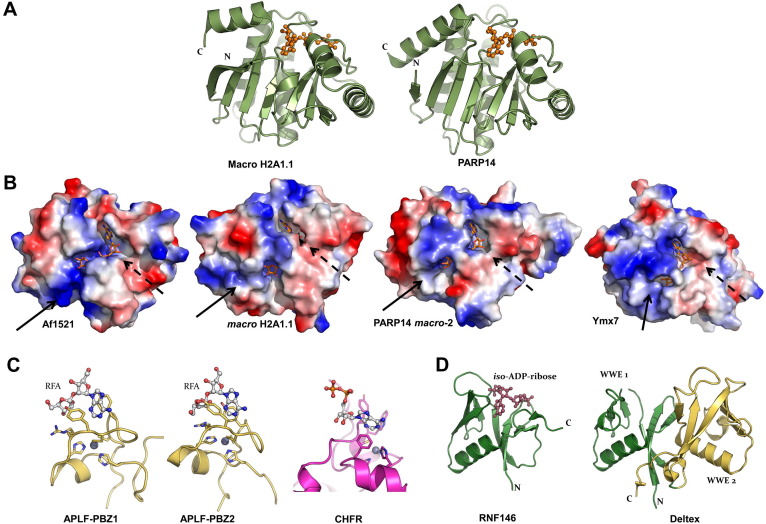
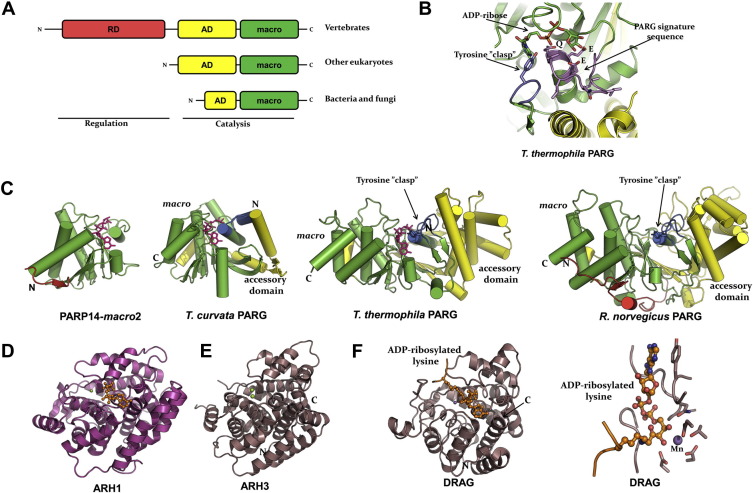
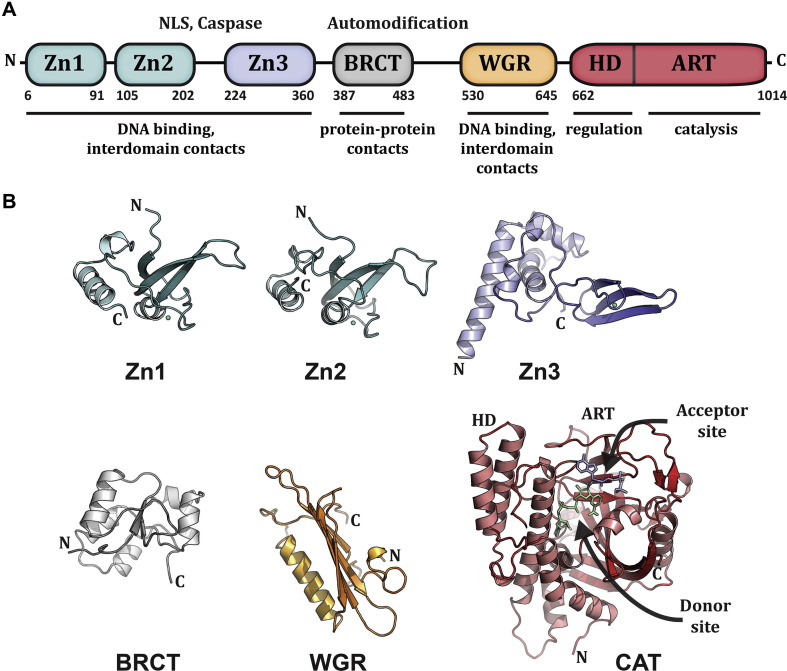
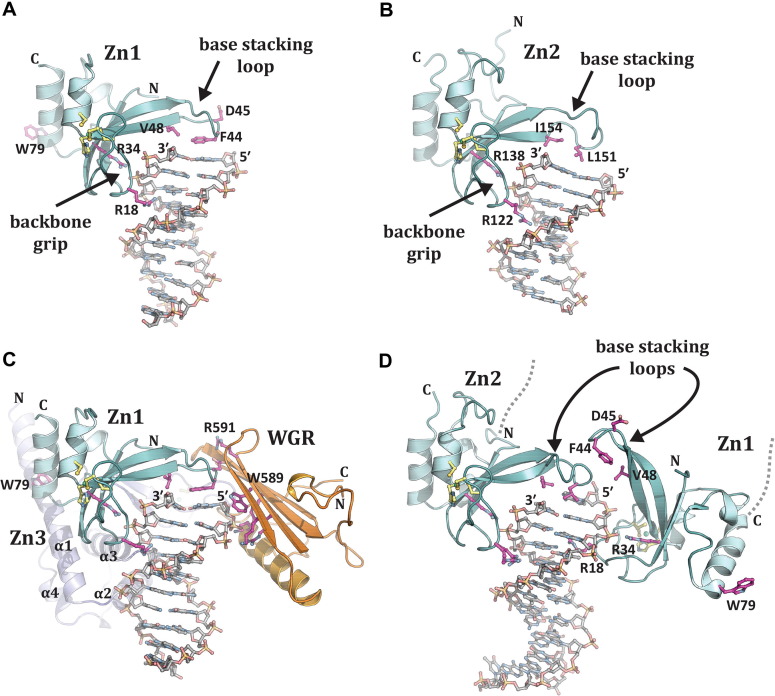
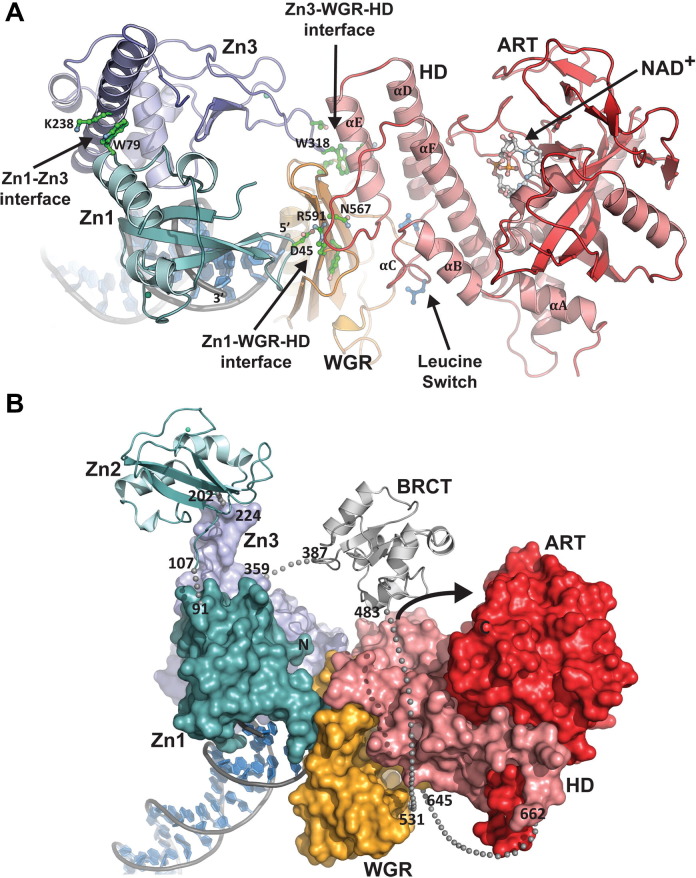
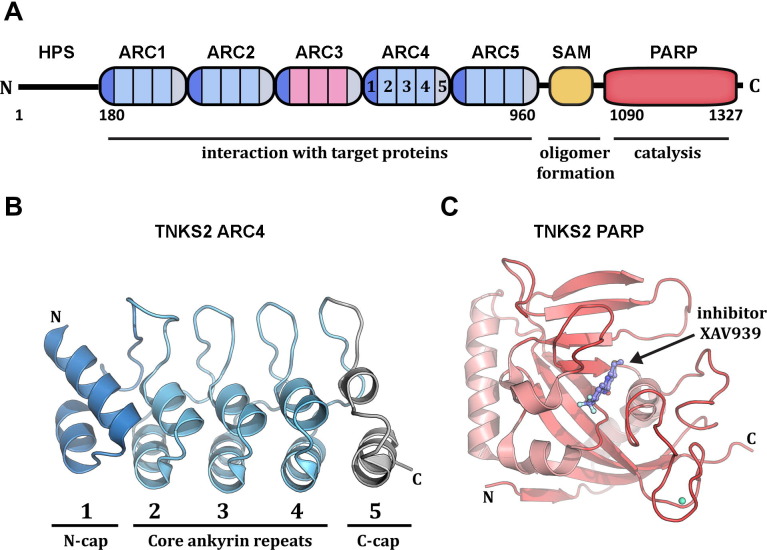
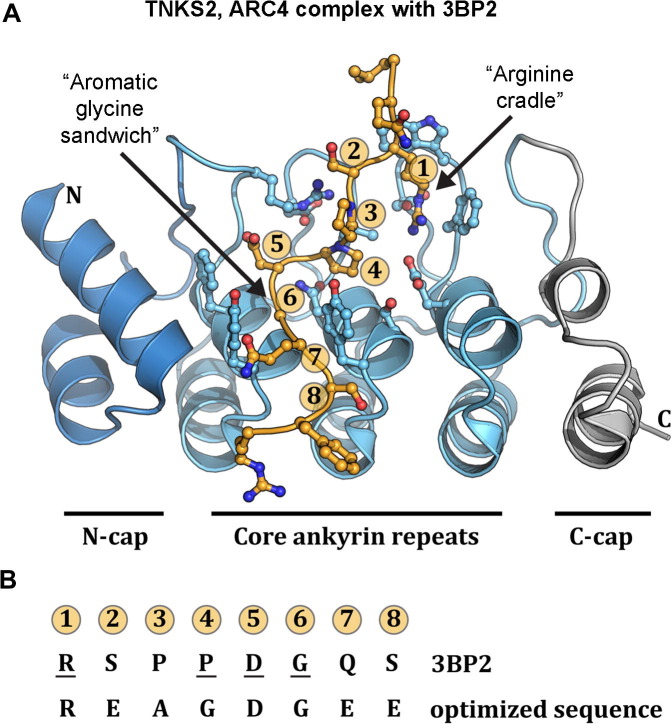
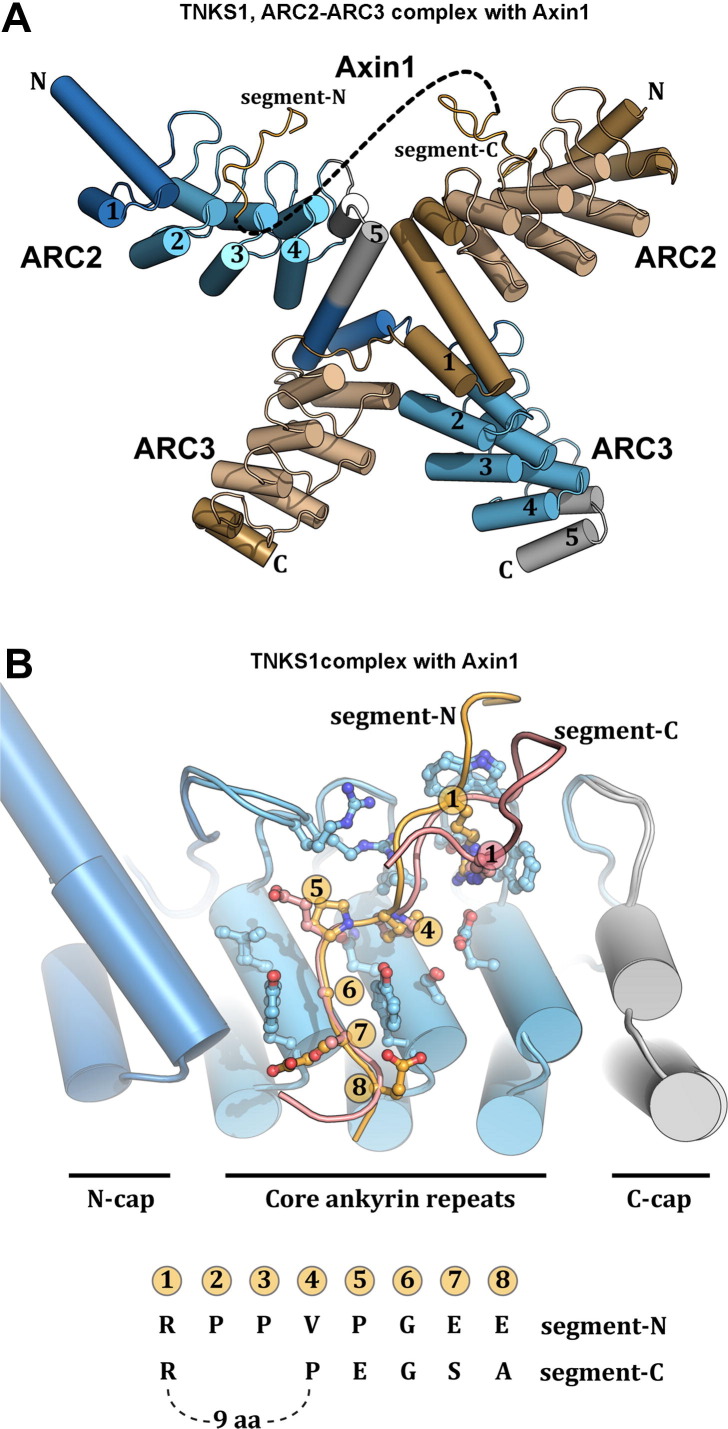
ADP-ribosylation of proteins regulates protein activities in various processes including transcription control, chromatin organization, organelle assembly, protein degradation, and DNA repair. Modulating the proteins involved in the metabolism of ADP-ribosylation can have therapeutic benefits in various disease states. Protein crystal structures can help understand the biological functions, facilitate detailed analysis of single residues, as well as provide a basis for development of small molecule effectors. Here we review recent advances in our understanding of the structural biology of the writers, readers, and erasers of ADP-ribosylation.
Keywords: ADP-ribosylation; DNA repair; Glycohydrolase; Macro domain; Signaling pathways; Transferase.
Copyright © 2013 Elsevier Ltd. All rights reserved.
Figures
References
-
- Ahel I., Ahel D., Matsusaka T., Clark A.J., Pines J., Boulton S.J., West S.C. Poly(ADP-ribose)-binding zinc finger motifs in DNA repair/checkpoint proteins. Nature. 2008;451:81–85. - PubMed
-
- Ahuja N., Schwer B., Carobbio S., Waltregny D., North B.J., Castronovo V., Maechler P., Verdin E. Regulation of insulin secretion by SIRT4, a mitochondrial ADP-ribosyltransferase. J. Biol. Chem. 2007;282:33583–33592. - PubMed
-
- Allen M.D., Buckle A.M., Cordell S.C., Lowe J., Bycroft M. The crystal structure of AF1521 a protein from Archaeoglobus fulgidus with homology to the non-histone domain of macroH2A. J. Mol. Biol. 2003;330:503–511. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
