

Long-term supplementation with a cystine-based antioxidant delays loss of muscle mass in aging
- PMID: 23459206
- PMCID: PMC3674717
- DOI: 10.1093/gerona/gls334
Long-term supplementation with a cystine-based antioxidant delays loss of muscle mass in aging
Abstract
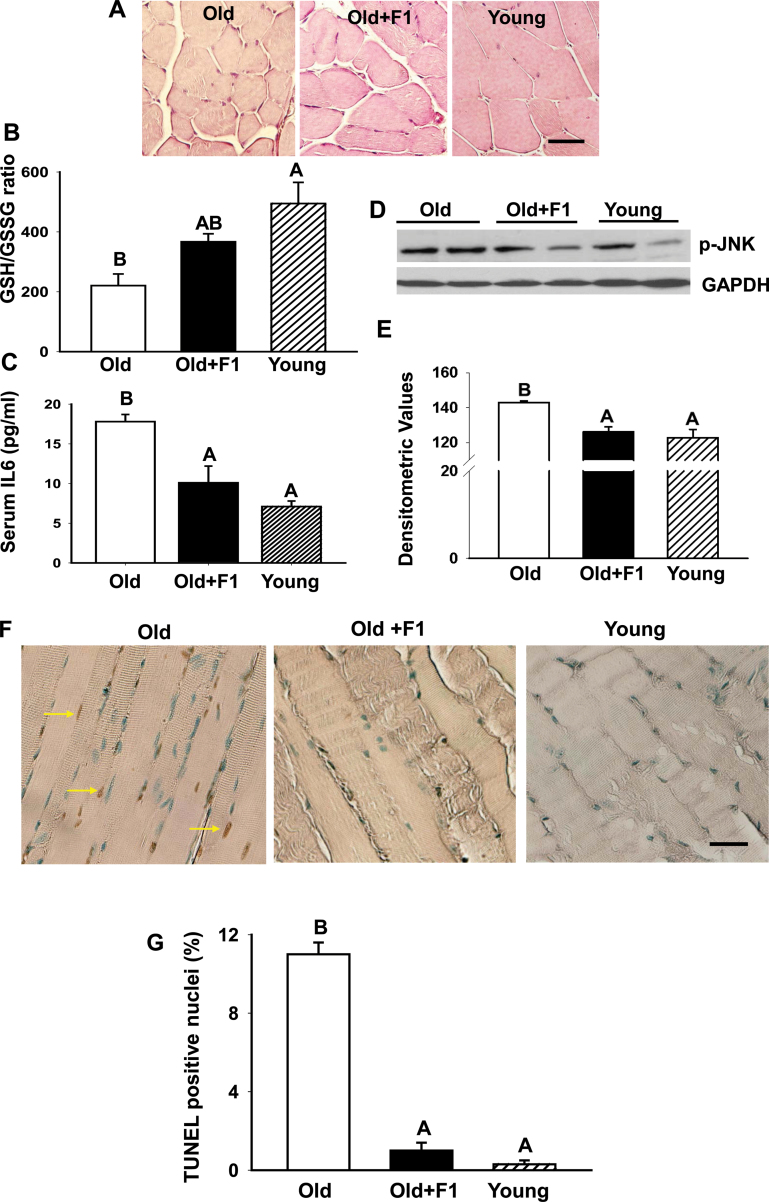
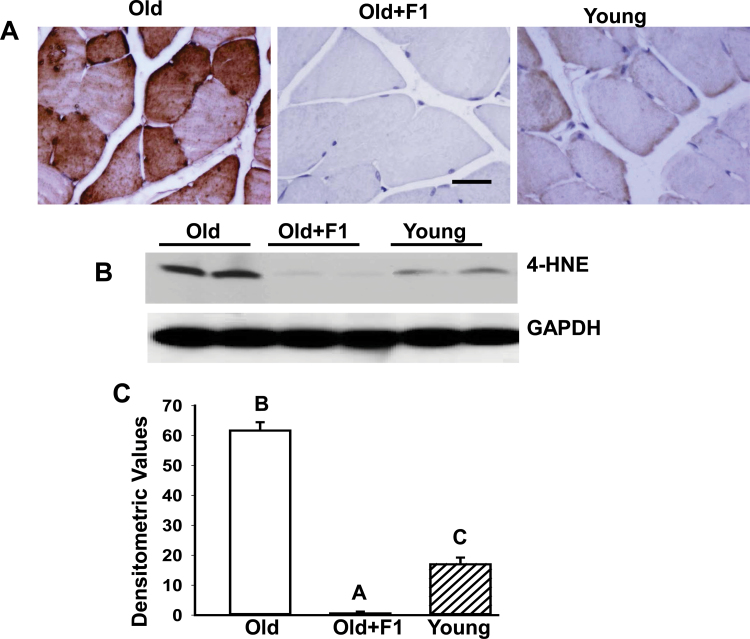
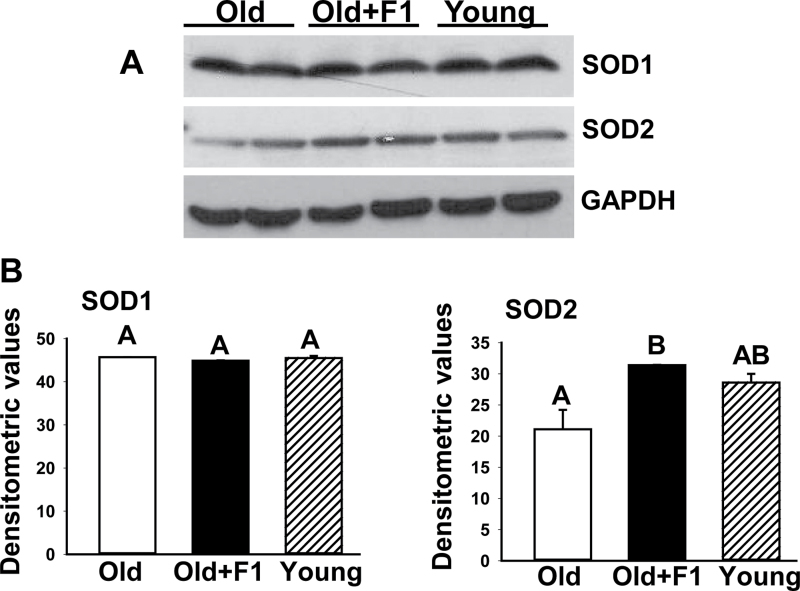
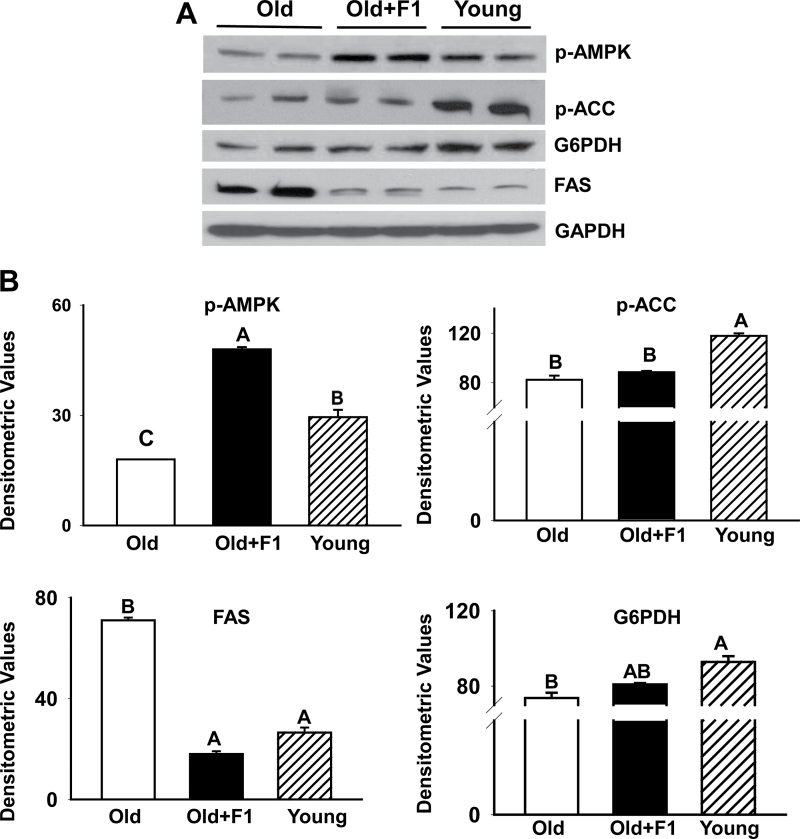
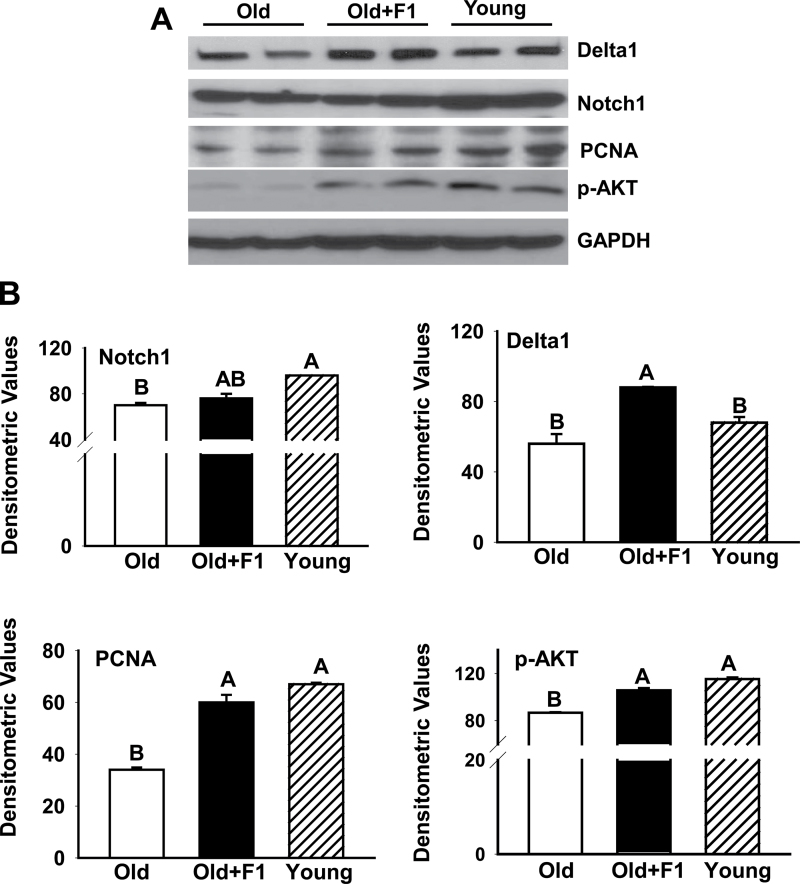
Oxidative stress increases with age and is postulated to be a major causal factor for sarcopenia in aging. Here, we examined whether the administration of a cystine-based antioxidant (F1) can alleviate/delay age-specific changes in skeletal muscles. C57BL6 male mice aged 17 months (middle aged) were fed with normal diet with or without supplementation of F1 (3 mg/kg food) for 6 months. Compared with young (5 months old) mice old mice exhibited increased markers of oxidative stress, inflammation, and muscle cell apoptosis and decreased muscle weight. These age-related changes were further associated with inactivation of adenosine-5'-monophosphate-activated protein kinase (AMPK), increased lipogenesis, activation of c-Jun NH2-terminal kinase, and decreased expression of Delta 1, phospho-Akt, and proliferating cell nuclear antigen in aged skeletal muscle. Such alterations were significantly prevented by F1. These results demonstrate the beneficial effects of F1 to attenuate loss of muscle mass associated with aging.
Keywords: Antioxidant; Mice; Muscle cell apoptosis; Oxidative stress; Sarcopenia..
Figures
Similar articles
-
Testosterone supplementation reverses sarcopenia in aging through regulation of myostatin, c-Jun NH2-terminal kinase, Notch, and Akt signaling pathways.Endocrinology. 2010 Feb;151(2):628-38. doi: 10.1210/en.2009-1177. Epub 2009 Dec 18. Endocrinology. 2010. PMID: 20022929 Free PMC article.
-
The histone deacetylase inhibitor butyrate improves metabolism and reduces muscle atrophy during aging.Aging Cell. 2015 Dec;14(6):957-70. doi: 10.1111/acel.12387. Epub 2015 Aug 20. Aging Cell. 2015. PMID: 26290460 Free PMC article.
-
Skeletal Muscle Cell Oxidative Stress as a Possible Therapeutic Target in a Denervation-Induced Experimental Sarcopenic Model.Spine (Phila Pa 1976). 2019 Apr 15;44(8):E446-E455. doi: 10.1097/BRS.0000000000002891. Spine (Phila Pa 1976). 2019. PMID: 30299418
-
Redox Systems, Antioxidants and Sarcopenia.Curr Protein Pept Sci. 2018;19(7):643-648. doi: 10.2174/1389203718666170317120040. Curr Protein Pept Sci. 2018. PMID: 28317484 Review.
-
Potential roles of vitamin E in age-related changes in skeletal muscle health.Nutr Res. 2018 Jan;49:23-36. doi: 10.1016/j.nutres.2017.09.005. Epub 2017 Sep 21. Nutr Res. 2018. PMID: 29420990 Review.
Cited by
-
Resveratrol encapsulated in novel fusogenic liposomes activates Nrf2 and attenuates oxidative stress in cerebromicrovascular endothelial cells from aged rats.J Gerontol A Biol Sci Med Sci. 2015 Mar;70(3):303-13. doi: 10.1093/gerona/glu029. Epub 2014 Mar 18. J Gerontol A Biol Sci Med Sci. 2015. PMID: 24642904 Free PMC article.
-
High whey protein intake delayed the loss of lean body mass in healthy old rats, whereas protein type and polyphenol/antioxidant supplementation had no effects.PLoS One. 2014 Sep 30;9(9):e109098. doi: 10.1371/journal.pone.0109098. eCollection 2014. PLoS One. 2014. PMID: 25268515 Free PMC article.
-
Comparative proteomic profiling reveals a role for Cisd2 in skeletal muscle aging.Aging Cell. 2018 Feb;17(1):e12705. doi: 10.1111/acel.12705. Epub 2017 Nov 23. Aging Cell. 2018. PMID: 29168286 Free PMC article.
-
Beneficial Effect of H2S-Releasing Molecules in an In Vitro Model of Sarcopenia: Relevance of Glucoraphanin.Int J Mol Sci. 2022 May 25;23(11):5955. doi: 10.3390/ijms23115955. Int J Mol Sci. 2022. PMID: 35682634 Free PMC article.
-
Parkin overexpression protects from ageing-related loss of muscle mass and strength.J Physiol. 2019 Apr;597(7):1975-1991. doi: 10.1113/JP277157. Epub 2019 Jan 30. J Physiol. 2019. PMID: 30614532 Free PMC article.
References
-
- Census briefs: the older population. Werner, CA: United States Census Bureau, US Department of Commerce; 2010.
-
- Hughes VA, Frontera WR, Roubenoff R, Evans WJ, Singh MA. Longitudinal changes in body composition in older men and women: role of body weight change and physical activity. Am J Clin Nutr. 2002; 76: 473–481 - PubMed
-
- Marzetti E, Leeuwenburgh C. Skeletal muscle apoptosis, sarcopenia and frailty at old age. Exp Gerontol. 2006; 41: 1234–1238 - PubMed
-
- Baumgartner RN, Koehler KM, Gallagher D, et al. Epidemiology of sarcopenia among the elderly in New Mexico. Am J Epidemiol. 1998; 147: 755–763 - PubMed
-
- Glass D, Roubenoff R. Recent advances in the biology and therapy of muscle wasting. Ann N Y Acad Sci. 2010; 1211: 25–36 - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Medical
Miscellaneous
