

The social fabric of the RNA degradosome
- PMID: 23459248
- PMCID: PMC3991390
- DOI: 10.1016/j.bbagrm.2013.02.011
The social fabric of the RNA degradosome
Abstract
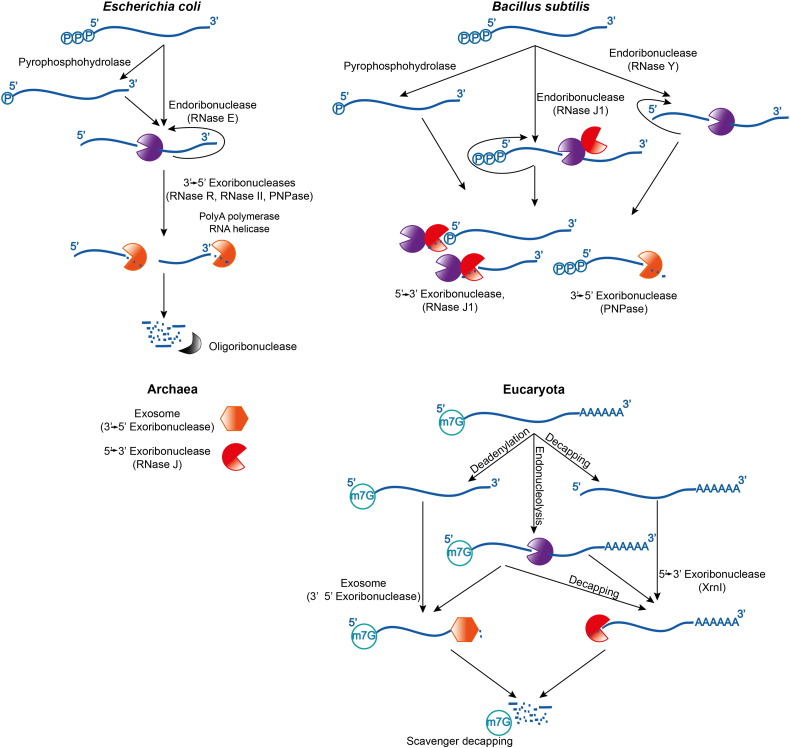
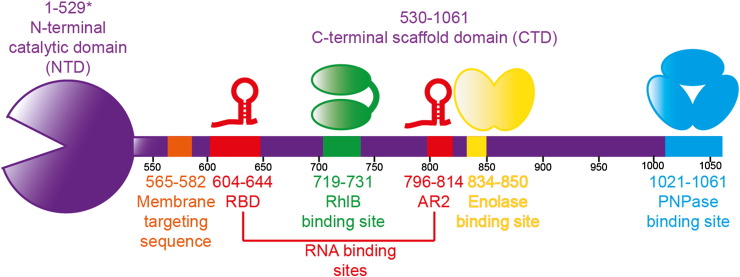
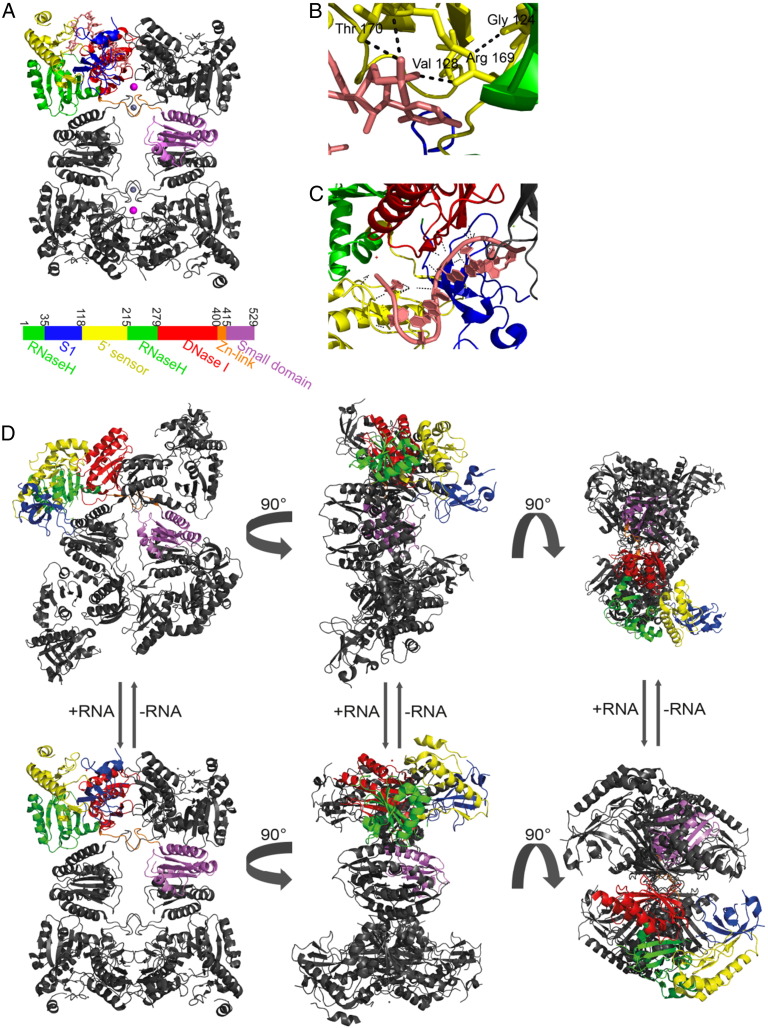
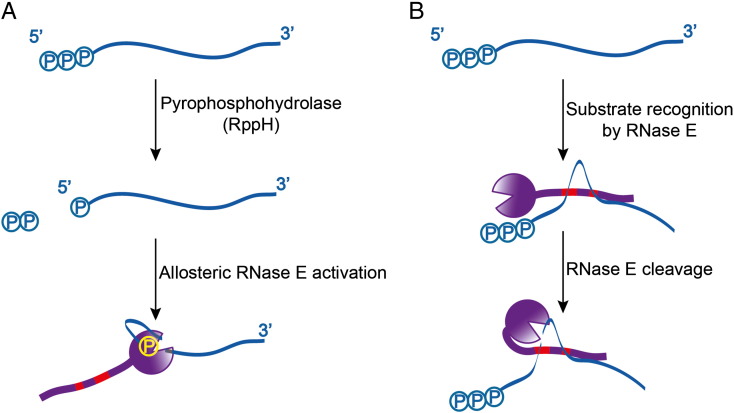
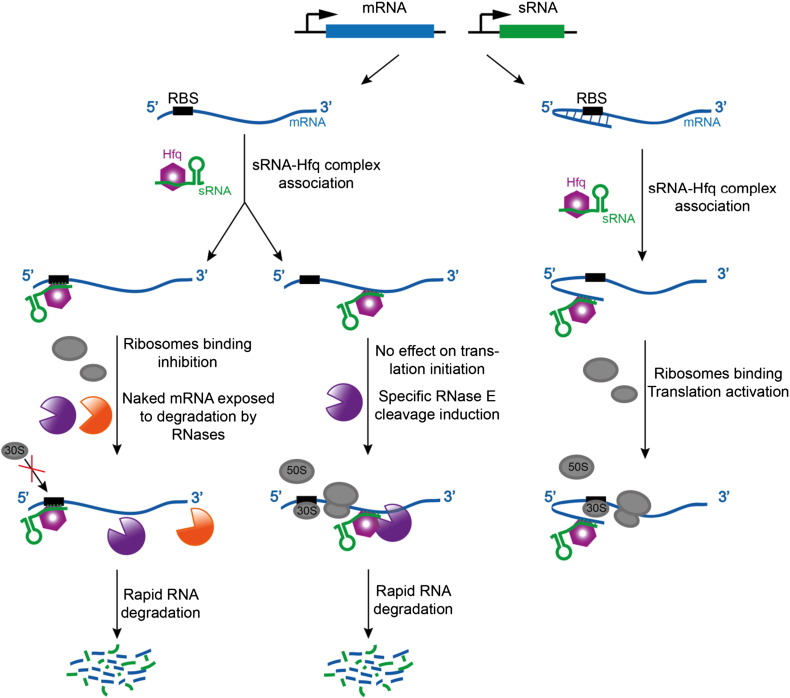
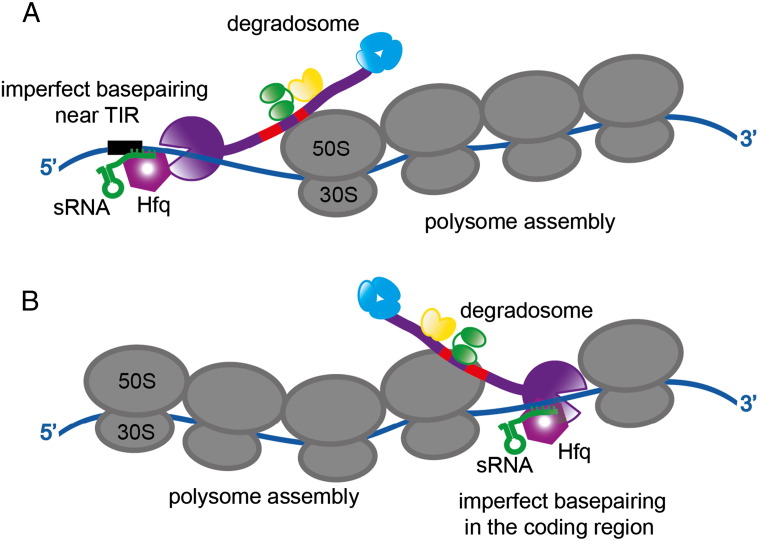
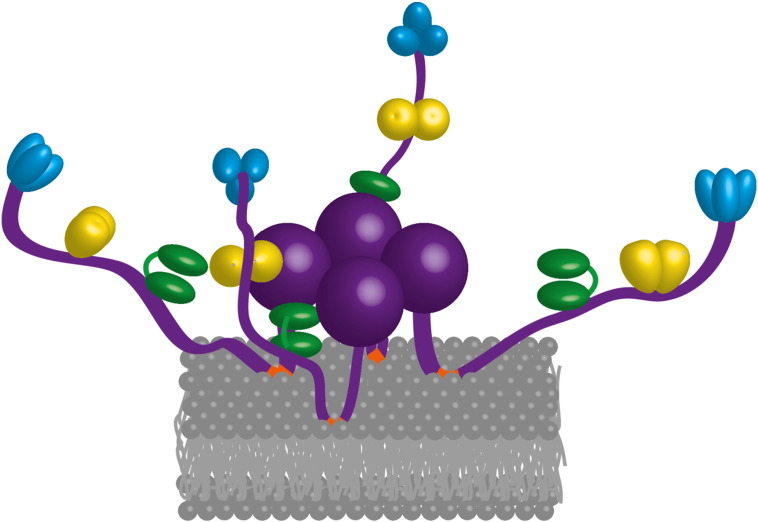
Bacterial transcripts each have a characteristic half-life, suggesting that the processes of RNA degradation work in an active and selective manner. Moreover, the processes are well controlled, thereby ensuring that degradation is orderly and coordinated. Throughout much of the bacterial kingdom, RNA degradation processes originate through the actions of assemblies of key RNA enzymes, known as RNA degradosomes. Neither conserved in composition, nor unified by common evolutionary ancestry, RNA degradosomes nonetheless can be found in divergent bacterial lineages, implicating a common requirement for the co-localisation of RNA metabolic activities. We describe how the cooperation of components in the representative degradosome of Escherichia coli may enable controlled access to transcripts, so that they have defined and programmable lifetimes. We also discuss how this cooperation contributes to precursor processing and to the riboregulation of intricate post-transcriptional networks in the control of gene expression. The E. coli degradosome interacts with the cytoplasmic membrane, and we discuss how this interaction may spatially organise the assembly and contribute to subunit cooperation and substrate capture. This article is part of a Special Issue entitled: RNA Decay mechanisms.
Copyright © 2013 Elsevier B.V. All rights reserved.
Figures
References
-
- Carpousis A.J. The RNA degradosome of Escherichia coli: an mRNA-degrading machine assembled on RNase E. Annu. Rev. Microbiol. 2007;61:71–87. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
