

Roux-en-Y gastric bypass reverses the effects of diet-induced obesity to inhibit the responsiveness of central vagal motoneurones
- PMID: 23459752
- PMCID: PMC3650700
- DOI: 10.1113/jphysiol.2012.249268
Roux-en-Y gastric bypass reverses the effects of diet-induced obesity to inhibit the responsiveness of central vagal motoneurones
Abstract
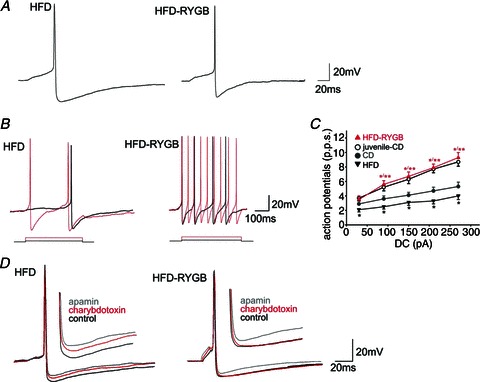
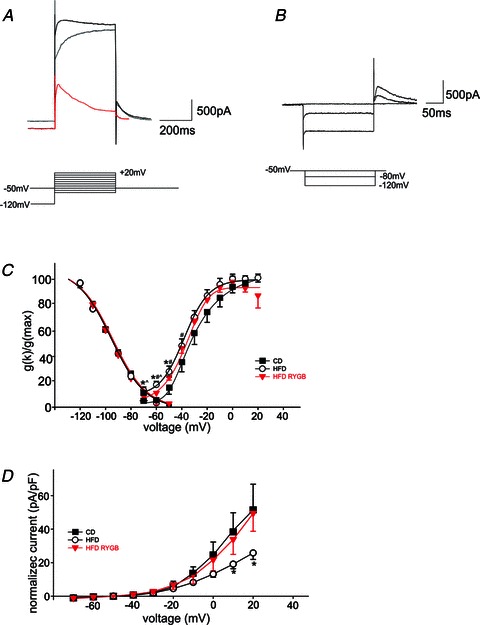
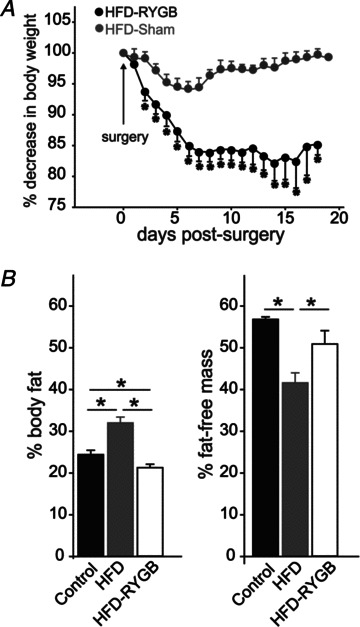
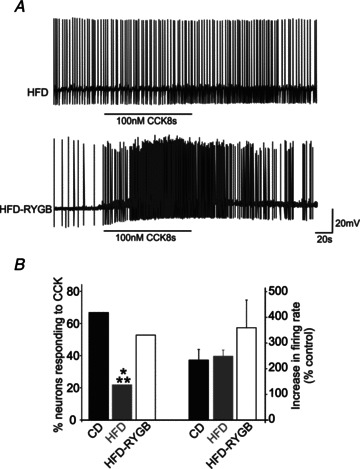
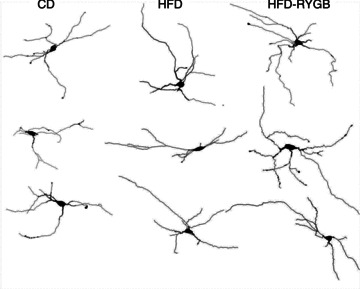
Diet-induced obesity (DIO) has been shown to alter the biophysical properties and pharmacological responsiveness of vagal afferent neurones and fibres, although the effects of DIO on central vagal neurones or vagal efferent functions have never been investigated. The aims of this study were to investigate whether high-fat diet-induced DIO also affects the properties of vagal efferent motoneurones, and to investigate whether these effects were reversed following weight loss induced by Roux-en-Y gastric bypass (RYGB) surgery. Whole-cell patch-clamp recordings were made from rat dorsal motor nucleus of the vagus (DMV) neurones in thin brainstem slices. The DMV neurones from rats exposed to high-fat diet for 12-14 weeks were less excitable, with a decreased membrane input resistance and decreased ability to fire action potentials in response to direct current pulse injection. The DMV neurones were also less responsive to superfusion with the satiety neuropeptides cholecystokinin and glucagon-like peptide 1. Roux-en-Y gastric bypass reversed all of these DIO-induced effects. Diet-induced obesity also affected the morphological properties of DMV neurones, increasing their size and dendritic arborization; RYGB did not reverse these morphological alterations. Remarkably, independent of diet, RYGB also reversed age-related changes of membrane properties and occurrence of charybdotoxin-sensitive (BK) calcium-dependent potassium current. These results demonstrate that DIO also affects the properties of central autonomic neurones by decreasing the membrane excitability and pharmacological responsiveness of central vagal motoneurones and that these changes were reversed following RYGB. In contrast, DIO-induced changes in morphological properties of DMV neurones were not reversed following gastric bypass surgery, suggesting that they may be due to diet, rather than obesity. These findings represent the first direct evidence for the plausible effect of RYGB to improve vagal neuronal health in the brain by reversing some effects of chronic high-fat diet as well as ageing. Vagovagal neurocircuits appear to remain open to modulation and adaptation throughout life, and understanding of these mechanisms may help in development of novel interventions to alleviate environmental (e.g. dietary) ailments and also alter neuronal ageing.
Figures
References
-
- Alywin ML, Horowitz JM, Bonham AC. NMDA receptors contribute to primary visceral afferent transmission in the nucleus of the solitary tract. J Neurophysiol. 1997;77:2539–2548. - PubMed
-
- Andresen MC, Yang M. Non-NMDA receptors mediate sensory afferent synaptic transmission in medial nucleus tractus solitarius. Am J Physiol Heart Circ Physiol. 1990;259:H1307–H1311. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Medical
Research Materials
Miscellaneous