

Review
doi: 10.1111/gtc.12040.
Epub 2013 Mar 5.
Loading and activation of DNA replicative helicases: the key step of initiation of DNA replication
Affiliations
- PMID: 23461534
- PMCID: PMC3657122
- DOI: 10.1111/gtc.12040
Item in Clipboard
Review
Loading and activation of DNA replicative helicases: the key step of initiation of DNA replication
Genes Cells.
2013 Apr.
Free PMC article
Abstract
Evolution has led to diversification of all living organisms from a common ancestor. Consequently, all living organisms use a common method to duplicate their genetic information and thus pass on their inherited traits to their offspring. To duplicate chromosomal DNA, double-stranded DNA must first be unwound by helicase, which is loaded to replication origins and activated during the DNA replication initiation step. In this review, we discuss the common features of, and differences in, replicative helicases between prokaryotes and eukaryotes.
© 2013 The Authors Genes to Cells © 2013 by the Molecular Biology Society of Japan and Wiley Publishing Asia Pty Ltd.
Figures
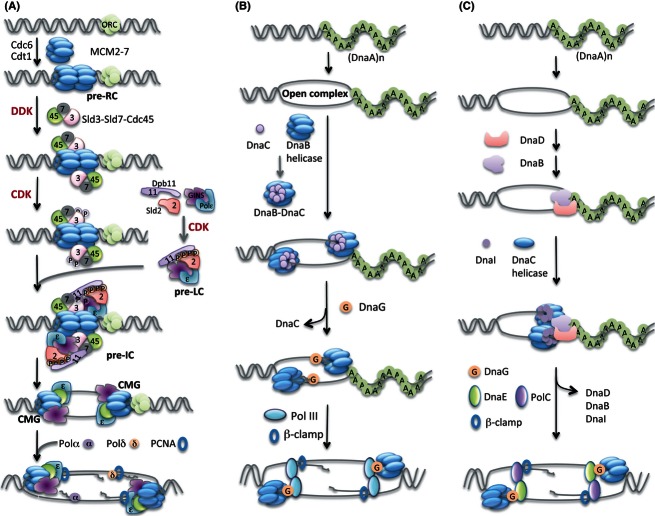
Origin loading and activation of replicative helicases. (A) In eukaryotes, the first step is loading of Mcm2–7 helicase onto the replication origin region that depends on origin recognition heterohexamer ORC and two loading proteins Cdc6 and Cdt1. In budding yeast S. cerevisiae, the subsequent activation of helicase requires DDK and CDK, recruiting other replication initiation proteins including a Sld3-Sld7-Cdc45 complex and then a preloading complex (pre-LC) composed of Sld2, Dpb11, GINS tetramer and polymerase DNA Pol ε onto the head-to-head double MCM hexamers to form a huge pre-initiation complex (pre-IC) at origin regions, immediately followed by remodeling that leads to the activation of CMG helicase. This process probably switches MCM from encircling dsDNA to ssDNA. Next, the active CMG works together with DNA polymerases, Pol α, Pol δ and Pol ε to duplicate DNA. (B) In Gram-negative E. coli, the initiator DnaA recognizes the bacterial replication origin oriC and melts a specific origin sequence to form an open complex in the presence of ATP. Then, DnaB–DnaC complex is loaded on each separated single-stranded DNA. The binding of primase DnaG with DnaB helicase and the primer synthesis induce removal of DnaC from DnaB. The DnaC dissociation requires ATPase activity. Subsequent loading of DNA replicative polymerase Pol III and β-clamp triggers replication elongation. (C) In Gram-positive B. subtilis, DnaA binds and melts oriC sequence, followed by loading of DnaD and DnaB orderly onto the origin region. DnaC helicase and DnaI assemble to the origin subsequently in the presence of ATP. The remodeling including removal of DnaD, DnaB and DnaI and the assembly of primase DnaG, two DNA polymerases, DnaE and PolC, and β-clamp activates helicase and replication fork movement.
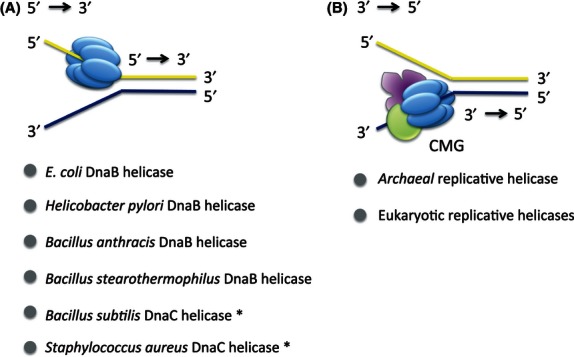
Unwinding polarity of replicative helicases. (A) Gram-negative bacteria E. coli and Helicobacter pylori helicase hexamer unwind double-stranded DNA in the direction of 5′ to 3′. Similarly, Gram-positive bacteria Bacillus anthracis and Bacillus stearothermophilus also showed a 5′ to 3′ unwinding activity. The DnaC helicase of Bacillus subtilis with 45% and 82% identical amino acid sequences to the counterpart in E. coli and Bacillus stearothermophilus, respectively, is speculated to have the same 5′ to 3′ helicase activity. Gram-positive Staphylococcus aureus DnaC helicase has 44% and 58% homology with E. coli DnaB and B. subtilis DnaC replicative DNA helicase, respectively, suggesting the same unwinding polarity as 5′ to 3′. (B) Both archaeal and eukaryotic CMG helicase complexes unwind double-stranded DNA in the direction 3′ to 5′.
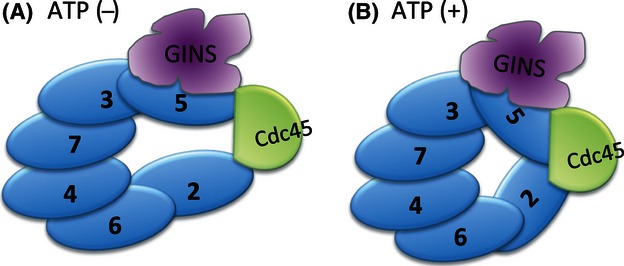
Conformational change of CMG. The CMG complex changes its conformation dynamically dependent on the presence of ATP. (A) In the absence of ATP, Mcm2 and Mcm5 disconnect and generate a big channel. (B) The binding of ATP leads to Mcm2 and Mcm5 closing the Mcm2–7 hexameric ring in assistance with Cdc45 and GINS.
References
-
- Allen GC, Jr, Kornberg A. Fine balance in the regulation of DnaB helicase by DnaC protein in replication in Escherichia coli. J. Biol. Chem. 1991;266:22096–22101. - PubMed
-
- Bell SP, Stillman B. ATP-dependent recognition of eukaryotic origins of DNA replication by a multiprotein complex. Nature. 1992;357:128–134. - PubMed
-
- Biswas SB, Biswas-Fiss EE. Quantitative analysis of binding of single-stranded DNA by Escherichia coli DnaB helicase and the DnaB x DnaC complex. Biochemistry. 2006;45:11505–11513. - PubMed
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Other Literature Sources
