

Coronary veins determine the pattern of sympathetic innervation in the developing heart
- PMID: 23462468
- PMCID: PMC3596991
- DOI: 10.1242/dev.087601
Coronary veins determine the pattern of sympathetic innervation in the developing heart
Abstract
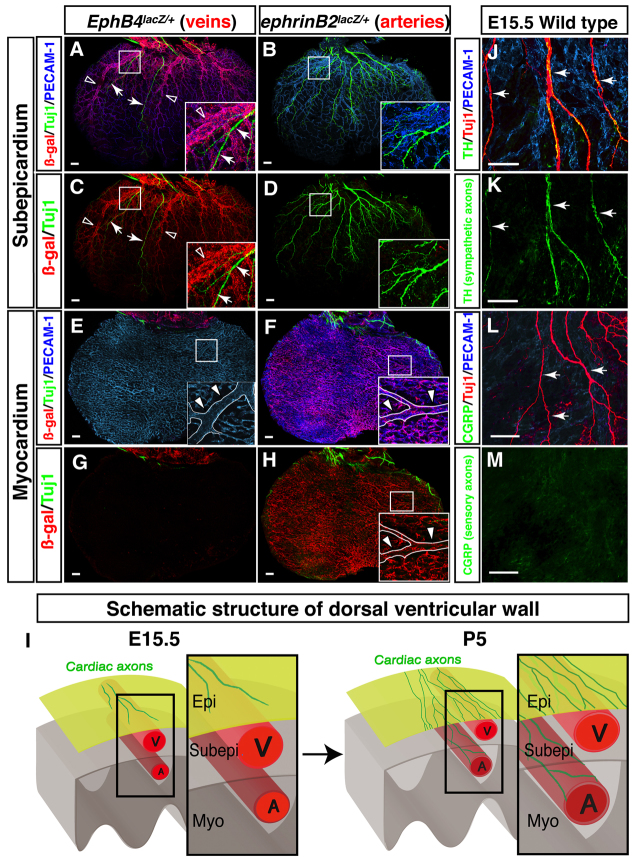
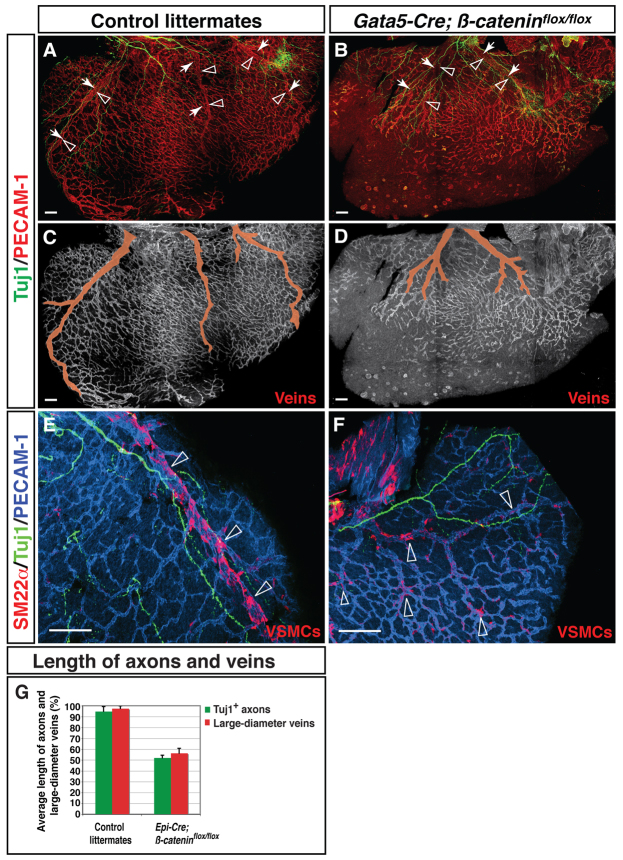
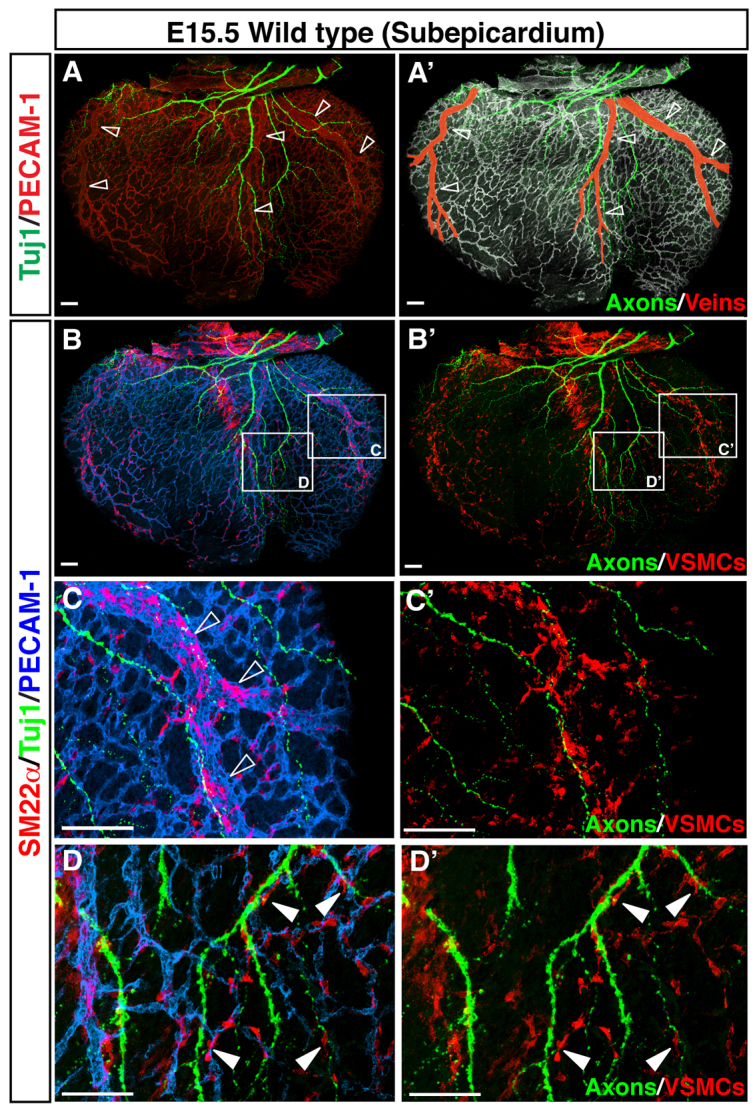
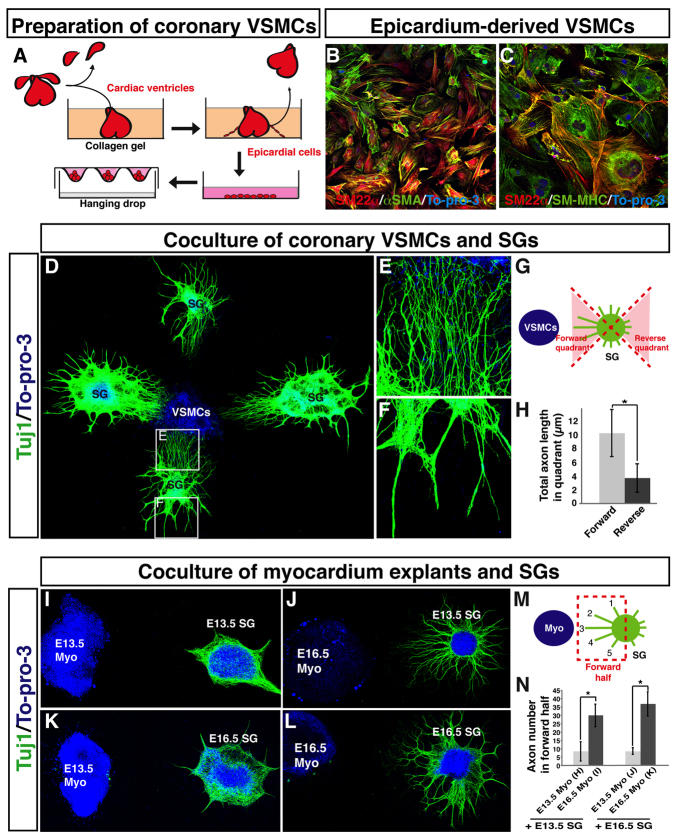
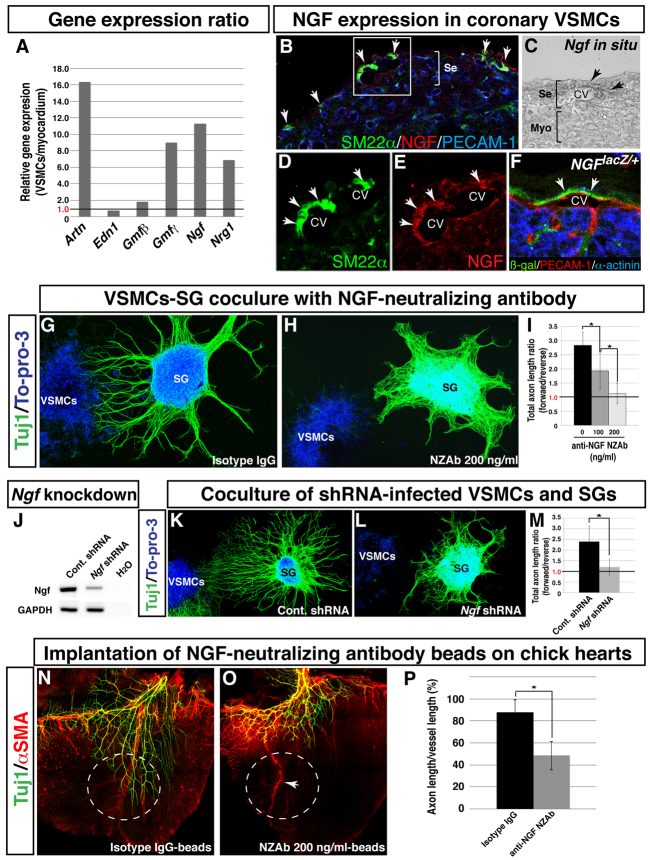
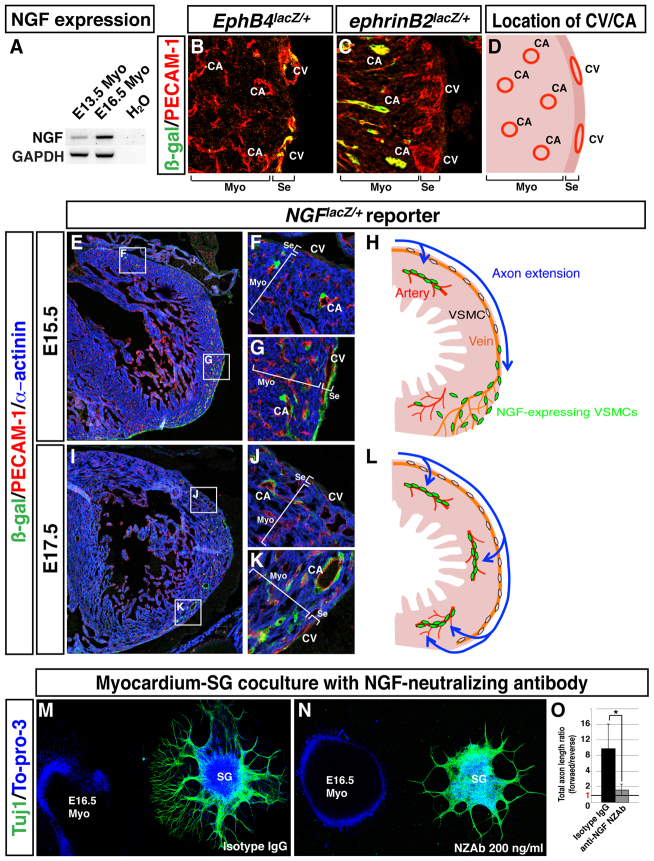
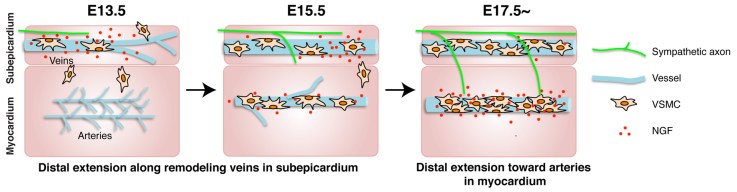
Anatomical congruence of peripheral nerves and blood vessels is well recognized in a variety of tissues. Their physical proximity and similar branching patterns suggest that the development of these networks might be a coordinated process. Here we show that large diameter coronary veins serve as an intermediate template for distal sympathetic axon extension in the subepicardial layer of the dorsal ventricular wall of the developing mouse heart. Vascular smooth muscle cells (VSMCs) associate with large diameter veins during angiogenesis. In vivo and in vitro experiments demonstrate that these cells mediate extension of sympathetic axons via nerve growth factor (NGF). This association enables topological targeting of axons to final targets such as large diameter coronary arteries in the deeper myocardial layer. As axons extend along veins, arterial VSMCs begin to secrete NGF, which allows axons to reach target cells. We propose a sequential mechanism in which initial axon extension in the subepicardium is governed by transient NGF expression by VSMCs as they are recruited to coronary veins; subsequently, VSMCs in the myocardium begin to express NGF as they are recruited by remodeling arteries, attracting axons toward their final targets. The proposed mechanism underlies a distinct, stereotypical pattern of autonomic innervation that is adapted to the complex tissue structure and physiology of the heart.
Figures

References
-
- Carmeliet P., Tessier-Lavigne M. (2005). Common mechanisms of nerve and blood vessel wiring. Nature 436, 193–200 - PubMed
-
- Enomoto H., Crawford P. A., Gorodinsky A., Heuckeroth R. O., Johnson E. M., Jr, Milbrandt J. (2001). RET signaling is essential for migration, axonal growth and axon guidance of developing sympathetic neurons. Development 128, 3963–3974 - PubMed
-
- Francis N., Farinas I., Brennan C., Rivas-Plata K., Backus C., Reichardt L., Landis S. (1999). NT-3, like NGF, is required for survival of sympathetic neurons, but not their precursors. Dev. Biol. 210, 411–427 - PubMed
Publication types
MeSH terms
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
