

The intracellular interactome of tetraspanin-enriched microdomains reveals their function as sorting machineries toward exosomes
- PMID: 23463506
- PMCID: PMC3636856
- DOI: 10.1074/jbc.M112.445304
The intracellular interactome of tetraspanin-enriched microdomains reveals their function as sorting machineries toward exosomes
Abstract
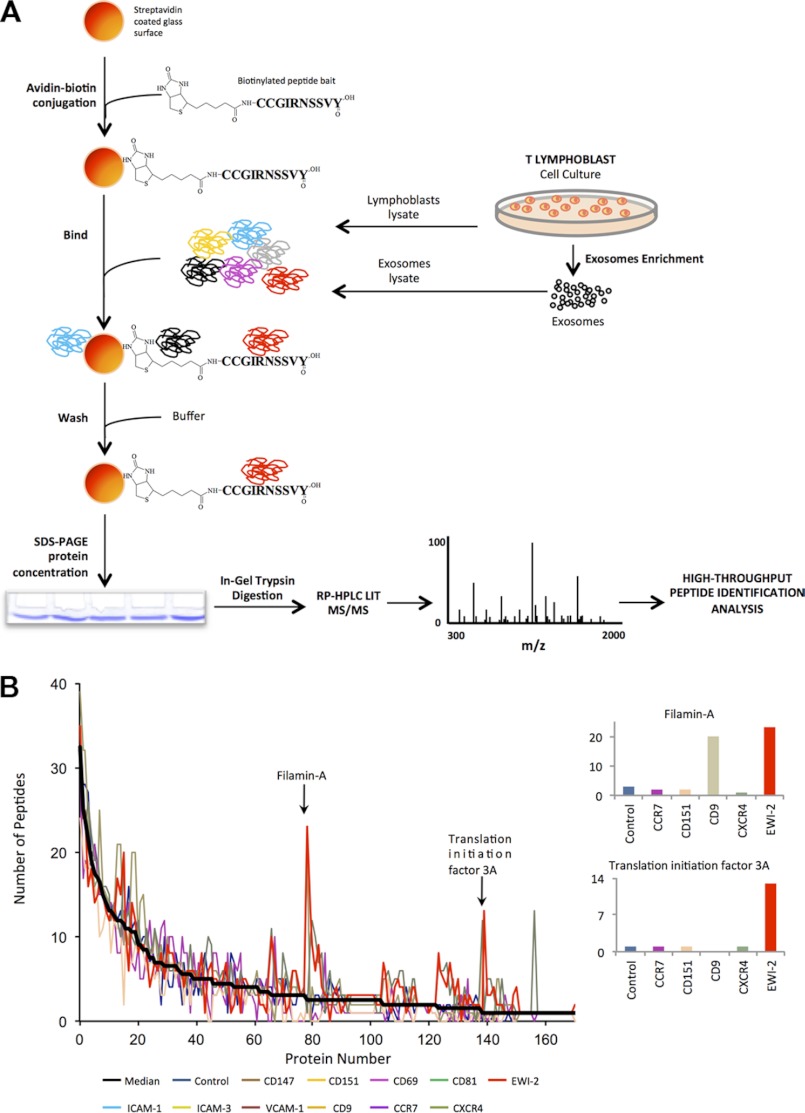
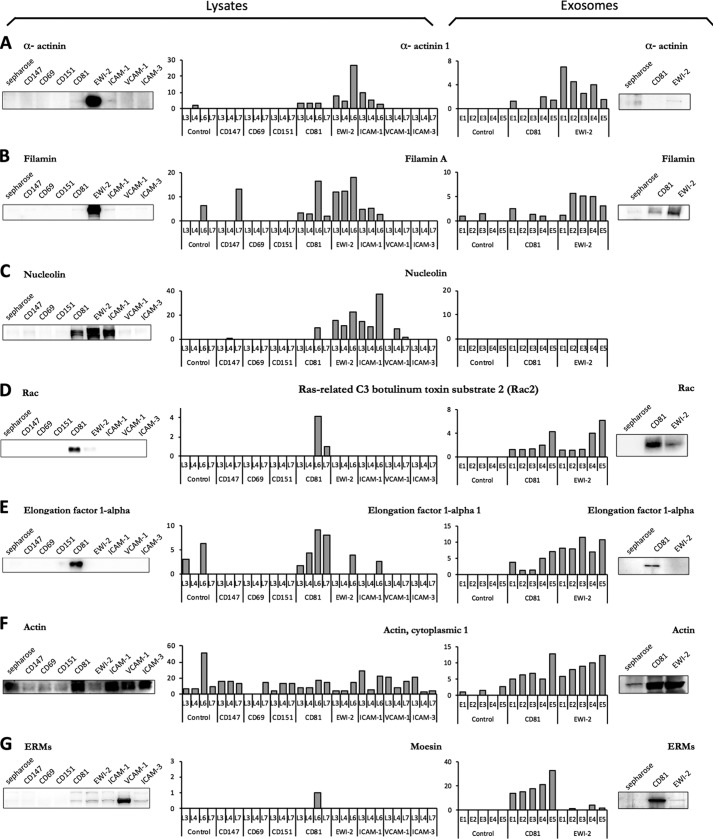
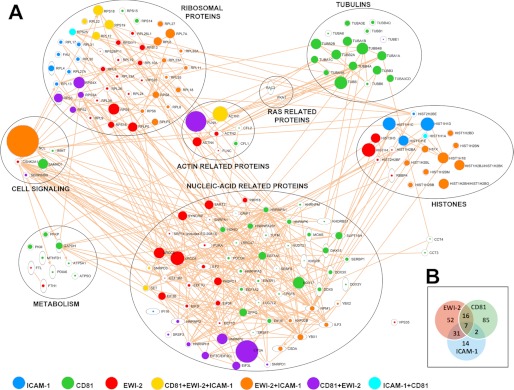
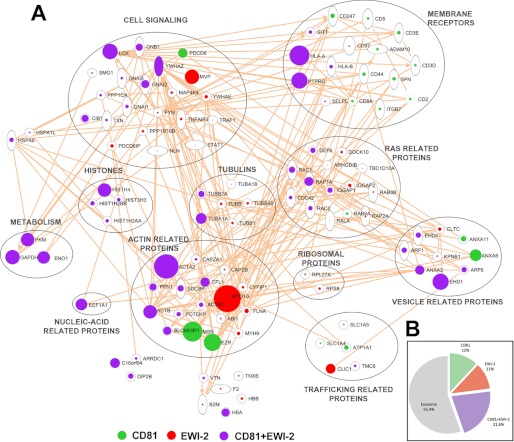
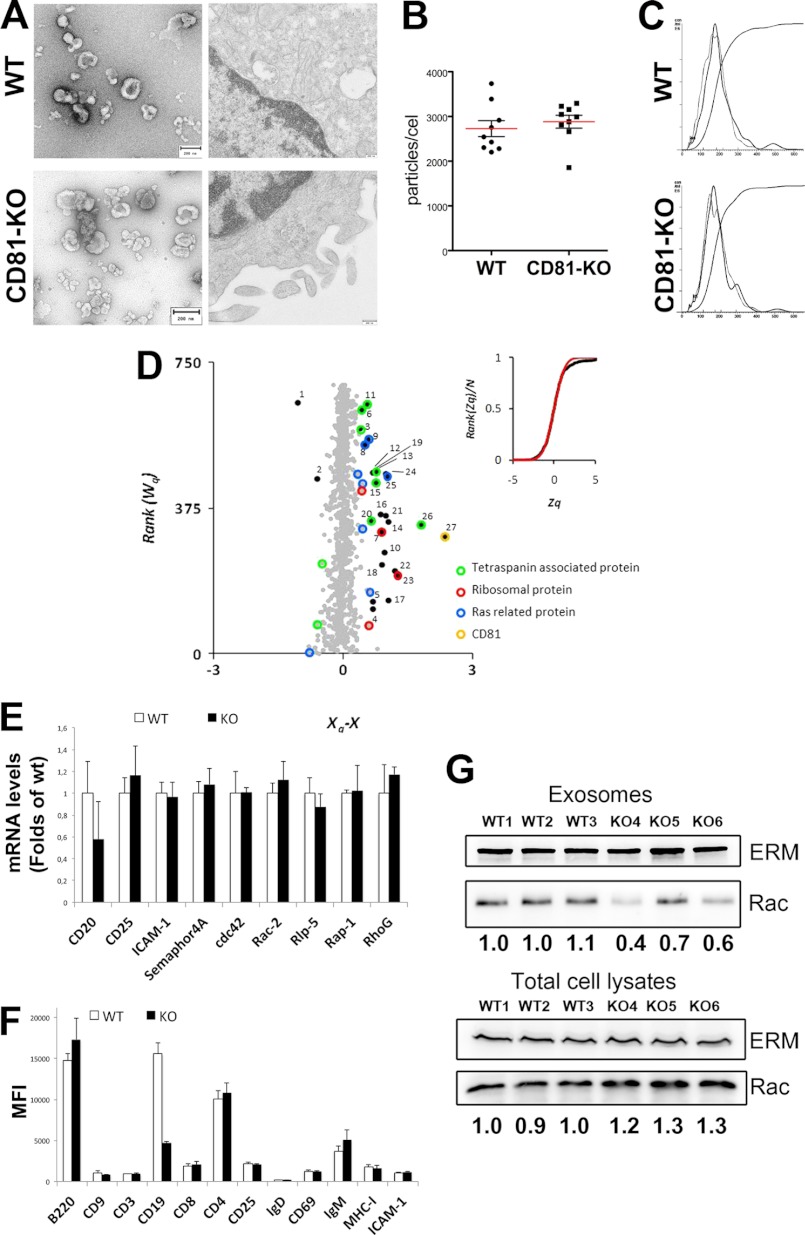
Extracellular vesicles are emerging as a potent mechanism of intercellular communication because they can systemically exchange genetic and protein material between cells. Tetraspanin molecules are commonly used as protein markers of extracellular vesicles, although their role in the unexplored mechanisms of cargo selection into exosomes has not been addressed. For that purpose, we have characterized the intracellular tetraspanin-enriched microdomain (TEM) interactome by high throughput mass spectrometry, in both human lymphoblasts and their derived exosomes, revealing a clear pattern of interaction networks. Proteins interacting with TEM receptors cytoplasmic regions presented a considerable degree of overlap, although some highly specific CD81 tetraspanin ligands, such as Rac GTPase, were detected. Quantitative proteomics showed that TEM ligands account for a great proportion of the exosome proteome and that a selective repertoire of CD81-associated molecules, including Rac, is not correctly routed to exosomes in cells from CD81-deficient animals. Our data provide evidence that insertion into TEM may be necessary for protein inclusion into the exosome structure.
Figures
References
-
- Yáñez-Mó M., Barreiro O., Gordon-Alonso M., Sala-Valdés M., Sánchez-Madrid F. (2009) Tetraspanin-enriched microdomains: a functional unit in cell plasma membranes. Trends Cell Biol. 19, 434–446 - PubMed
-
- Charrin S., le Naour F., Silvie O., Milhiet P. E., Boucheix C., Rubinstein E. (2009) Lateral organization of membrane proteins: tetraspanins spin their web. Biochem. J. 420, 133–154 - PubMed
-
- Berditchevski F., Odintsova E. (2007) Tetraspanins as regulators of protein trafficking. Traffic 8, 89–96 - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Miscellaneous
