

Thylakoid terminal oxidases are essential for the cyanobacterium Synechocystis sp. PCC 6803 to survive rapidly changing light intensities
- PMID: 23463783
- PMCID: PMC3641225
- DOI: 10.1104/pp.112.210260
Thylakoid terminal oxidases are essential for the cyanobacterium Synechocystis sp. PCC 6803 to survive rapidly changing light intensities
Abstract
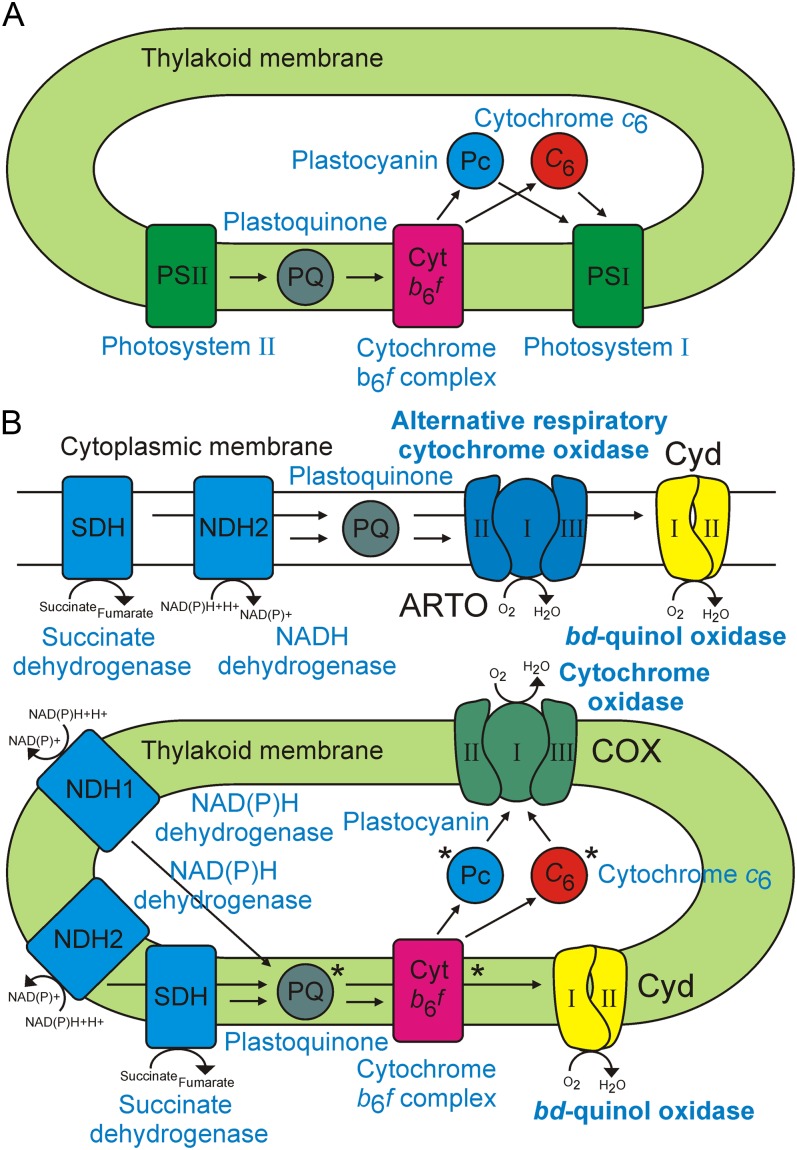
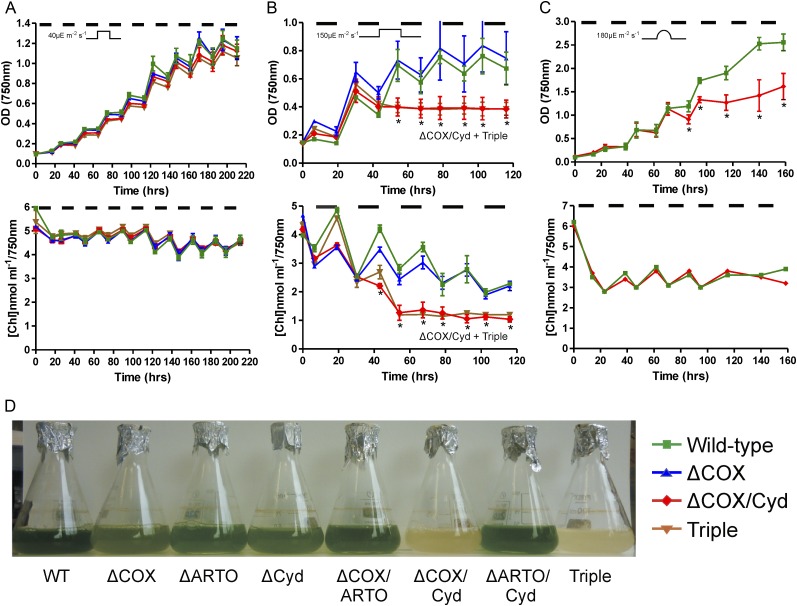
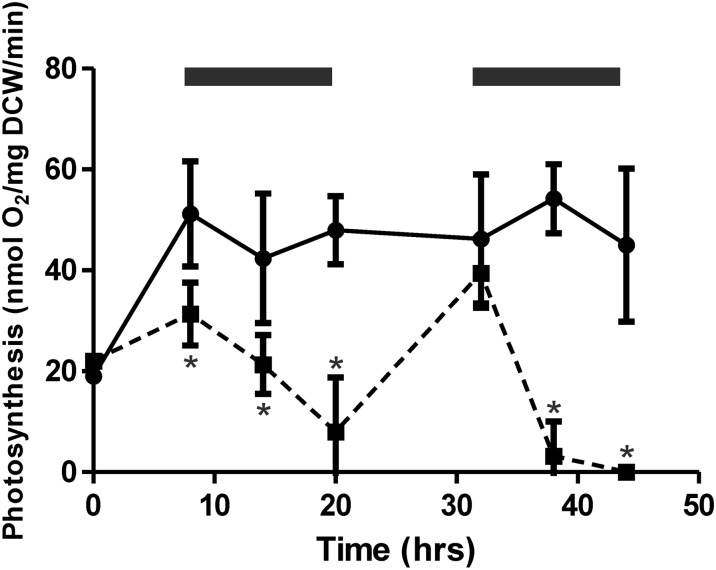
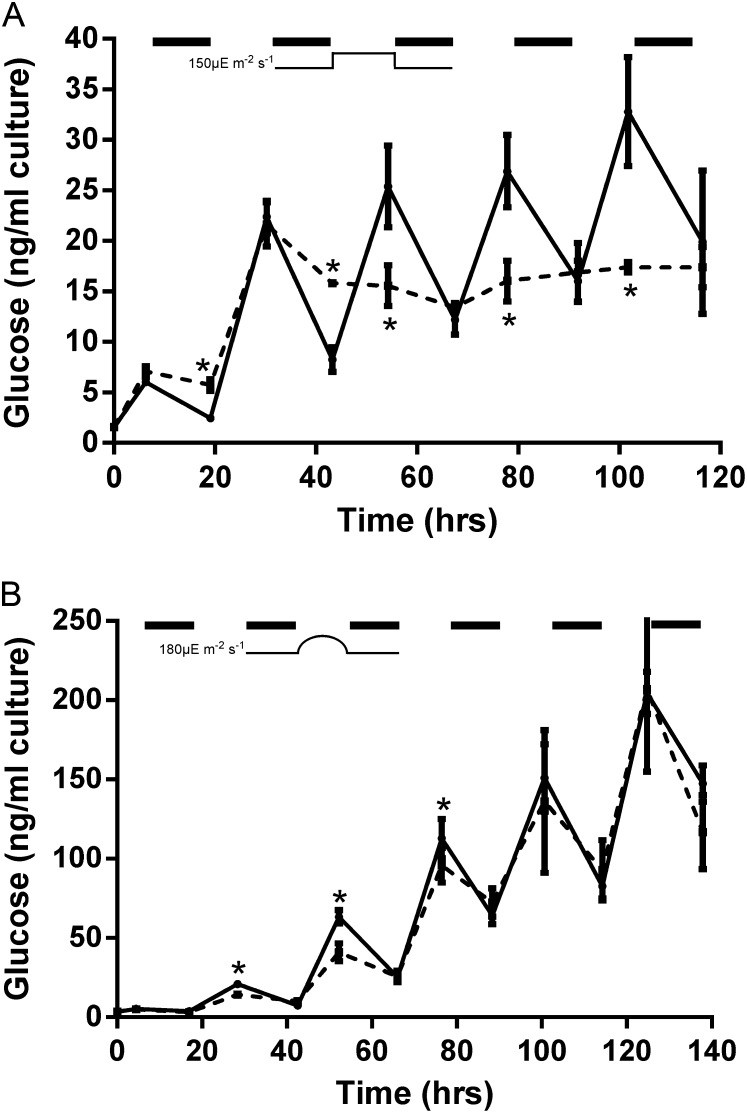
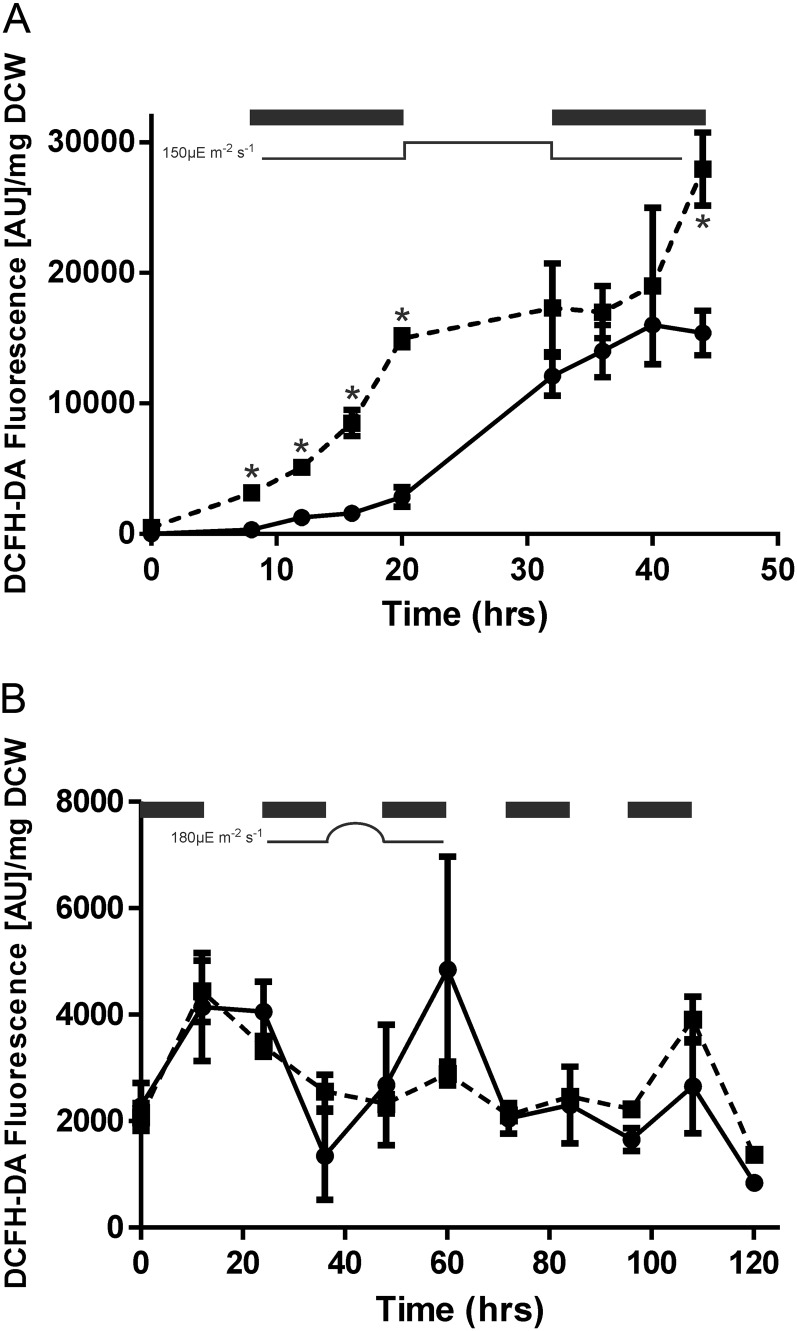
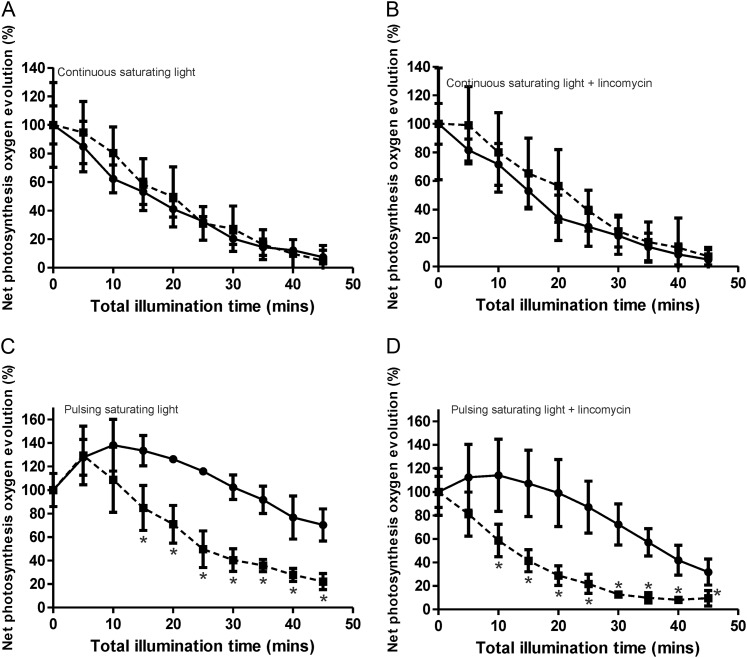
Cyanobacteria perform photosynthesis and respiration in the thylakoid membrane, suggesting that the two processes are interlinked. However, the role of the respiratory electron transfer chain under natural environmental conditions has not been established. Through targeted gene disruption, mutants of Synechocystis sp. PCC 6803 were generated that lacked combinations of the three terminal oxidases: the thylakoid membrane-localized cytochrome c oxidase (COX) and quinol oxidase (Cyd) and the cytoplasmic membrane-localized alternative respiratory terminal oxidase. All strains demonstrated similar growth under continuous moderate or high light or 12-h moderate-light/dark square-wave cycles. However, under 12-h high-light/dark square-wave cycles, the COX/Cyd mutant displayed impaired growth and was completely photobleached after approximately 2 d. In contrast, use of sinusoidal light/dark cycles to simulate natural diurnal conditions resulted in little photobleaching, although growth was slower. Under high-light/dark square-wave cycles, the COX/Cyd mutant suffered a significant loss of photosynthetic efficiency during dark periods, a greater level of oxidative stress, and reduced glycogen degradation compared with the wild type. The mutant was susceptible to photoinhibition under pulsing but not constant light. These findings confirm a role for thylakoid-localized terminal oxidases in efficient dark respiration, reduction of oxidative stress, and accommodation of sudden light changes, demonstrating the strong selective pressure to maintain linked photosynthetic and respiratory electron chains within the thylakoid membrane. To our knowledge, this study is the first to report a phenotypic difference in growth between terminal oxidase mutants and wild-type cells and highlights the need to examine mutant phenotypes under a range of conditions.
Figures
References
-
- Abramson J, Riistama S, Larsson G, Jasaitis A, Svensson-Ek M, Laakkonen L, Puustinen A, Iwata S, Wikström M. (2000) The structure of the ubiquinol oxidase from Escherichia coli and its ubiquinone binding site. Nat Struct Biol 7: 910–917 - PubMed
-
- Agarwal R, Matros A, Melzer M, Mock HP, Sainis JK. (2010) Heterogeneity in thylakoid membrane proteome of Synechocystis 6803. J Proteomics 73: 976–991 - PubMed
-
- Altschul SF, Gish W, Miller W, Myers EW, Lipman DJ. (1990) Basic local alignment search tool. J Mol Biol 215: 403–410 - PubMed
-
- Bailey S, Melis A, Mackey KR, Cardol P, Finazzi G, van Dijken G, Berg GM, Arrigo K, Shrager J, Grossman A. (2008) Alternative photosynthetic electron flow to oxygen in marine Synechococcus. Biochim Biophys Acta 1777: 269–276 - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
