

CALHM1 ion channel mediates purinergic neurotransmission of sweet, bitter and umami tastes
- PMID: 23467090
- PMCID: PMC3600154
- DOI: 10.1038/nature11906
CALHM1 ion channel mediates purinergic neurotransmission of sweet, bitter and umami tastes
Abstract
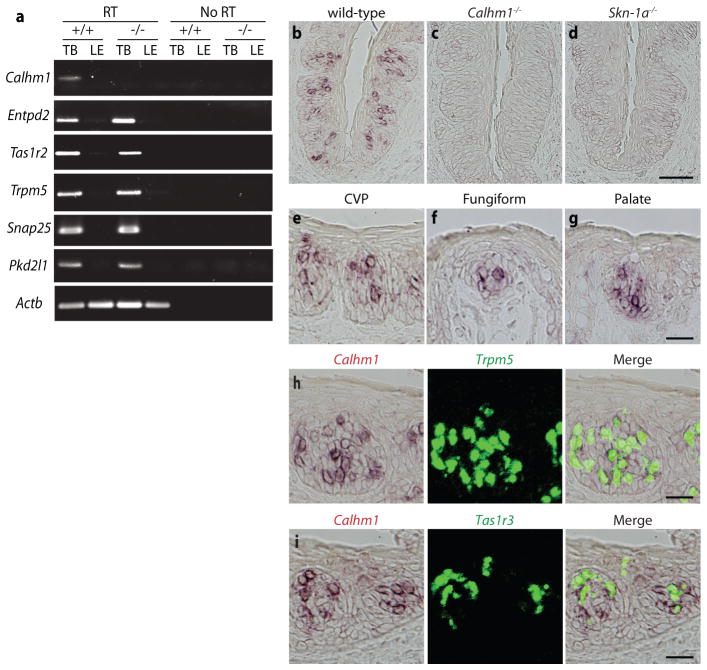
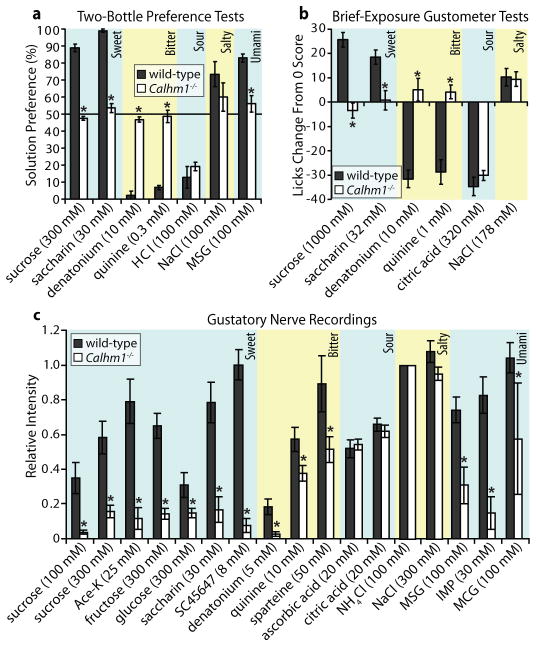
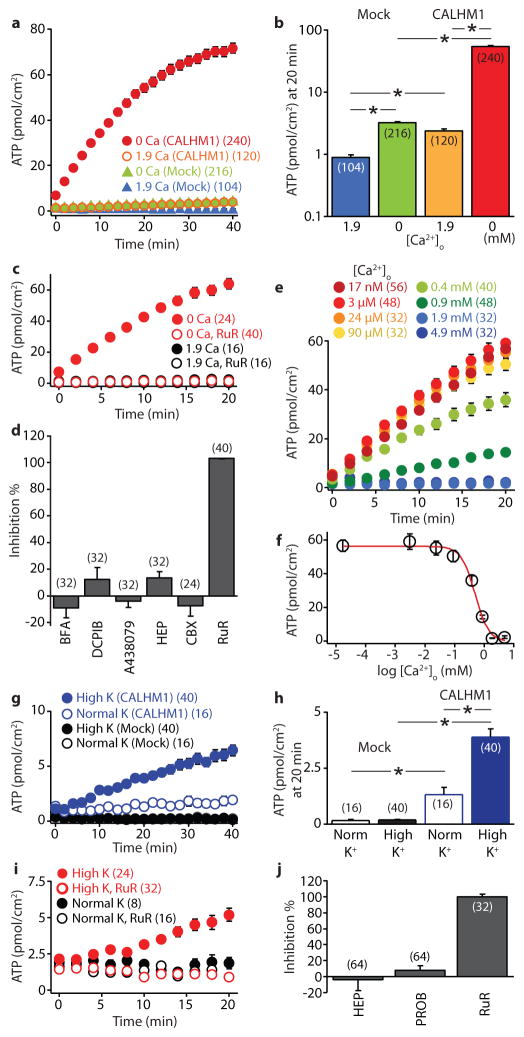
Recognition of sweet, bitter and umami tastes requires the non-vesicular release from taste bud cells of ATP, which acts as a neurotransmitter to activate afferent neural gustatory pathways. However, how ATP is released to fulfil this function is not fully understood. Here we show that calcium homeostasis modulator 1 (CALHM1), a voltage-gated ion channel, is indispensable for taste-stimuli-evoked ATP release from sweet-, bitter- and umami-sensing taste bud cells. Calhm1 knockout mice have severely impaired perceptions of sweet, bitter and umami compounds, whereas their recognition of sour and salty tastes remains mostly normal. Calhm1 deficiency affects taste perception without interfering with taste cell development or integrity. CALHM1 is expressed specifically in sweet/bitter/umami-sensing type II taste bud cells. Its heterologous expression induces a novel ATP permeability that releases ATP from cells in response to manipulations that activate the CALHM1 ion channel. Knockout of Calhm1 strongly reduces voltage-gated currents in type II cells and taste-evoked ATP release from taste buds without affecting the excitability of taste cells by taste stimuli. Thus, CALHM1 is a voltage-gated ATP-release channel required for sweet, bitter and umami taste perception.
Figures

References
References for Methods
Publication types
MeSH terms
Substances
Grants and funding
- R01 DC006308/DC/NIDCD NIH HHS/United States
- P30DC011735/DC/NIDCD NIH HHS/United States
- P30 DC011735/DC/NIDCD NIH HHS/United States
- R01 EY013624/EY/NEI NIH HHS/United States
- MH059937/MH/NIMH NIH HHS/United States
- GM56328/GM/NIGMS NIH HHS/United States
- R37 GM056328/GM/NIGMS NIH HHS/United States
- R01 DC010393/DC/NIDCD NIH HHS/United States
- R03DC011143/DC/NIDCD NIH HHS/United States
- R21 NS072775/NS/NINDS NIH HHS/United States
- NS072775/NS/NINDS NIH HHS/United States
- R01 GM056328/GM/NIGMS NIH HHS/United States
- R03 DC011143/DC/NIDCD NIH HHS/United States
- DC10393/DC/NIDCD NIH HHS/United States
- EY13624/EY/NEI NIH HHS/United States
- P30 EY001583/EY/NEI NIH HHS/United States
- R01 MH059937/MH/NIMH NIH HHS/United States
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
Research Materials
