

Adenosine transporter ENT1 regulates the acquisition of goal-directed behavior and ethanol drinking through A2A receptor in the dorsomedial striatum
- PMID: 23467349
- PMCID: PMC3622260
- DOI: 10.1523/JNEUROSCI.3094-12.2013
Adenosine transporter ENT1 regulates the acquisition of goal-directed behavior and ethanol drinking through A2A receptor in the dorsomedial striatum
Abstract
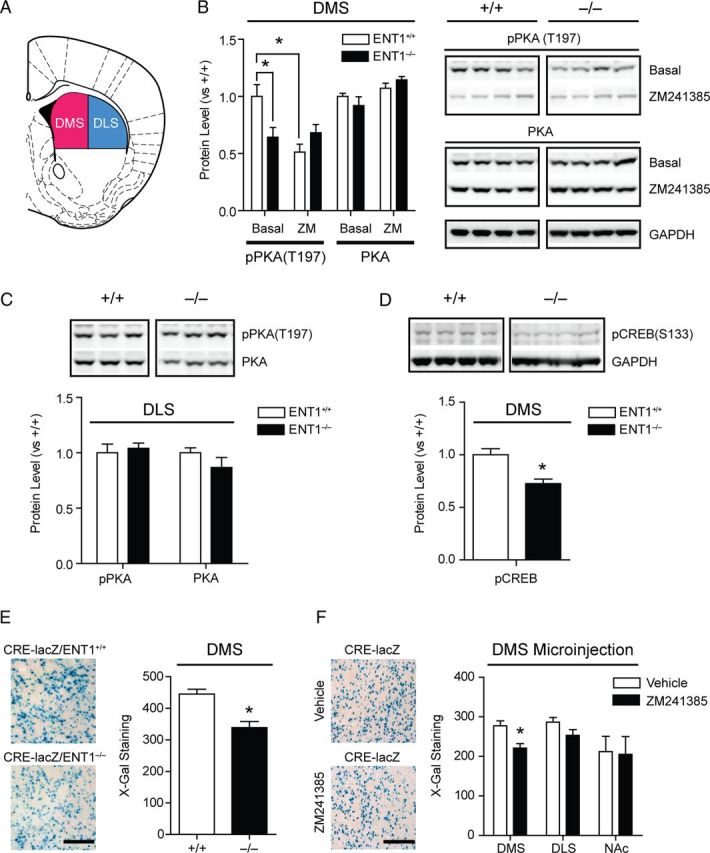
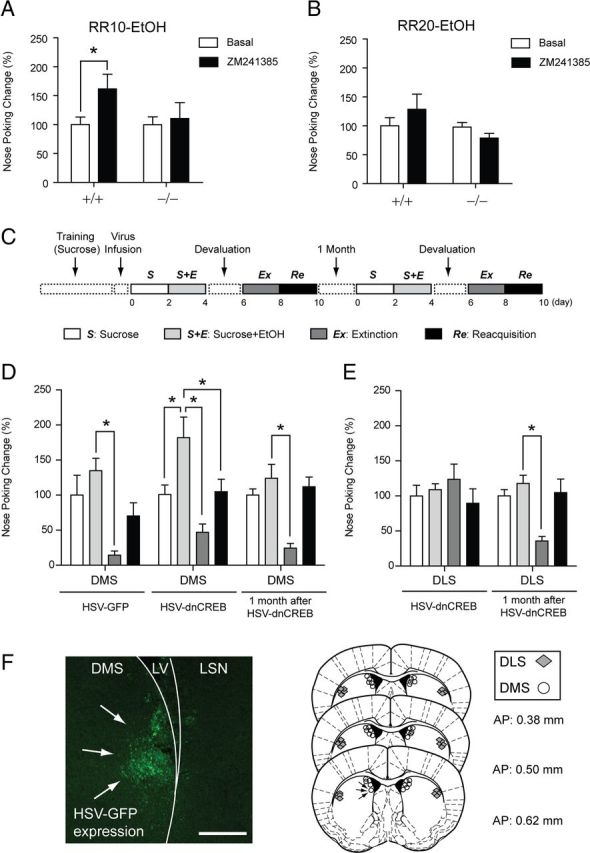
Adenosine signaling has been implicated in the pathophysiology of many psychiatric disorders including alcoholism. Striatal adenosine A2A receptors (A2AR) play an essential role in both ethanol drinking and the shift from goal-directed action to habitual behavior. However, direct evidence for a role of striatal A2AR signaling in ethanol drinking and habit development has not been established. In the present study, we found that decreased A2AR-mediated CREB activity in the dorsomedial striatum (DMS) enhanced initial behavioral acquisition of goal-directed behaviors and the vulnerability to progress to excessive ethanol drinking during operant conditioning in mice lacking ethanol-sensitive adenosine transporter ENT1 (ENT1(-/-)). Using mice expressing β-galactosidase (lacZ) under the control of seven repeated CRE sites in both genotypes (CRE-lacZ/ENT1(+/+) mice and CRE-lacZ/ENT1(-/-) mice) and the dominant-negative form of CREB, we found that reduced CREB activity in the DMS was causally associated with decreased A2AR signaling and increased goal-directed ethanol drinking. Finally, we have demonstrated that the A2AR antagonist ZM241385 dampened protein kinase A activity-mediated signaling in the DMS and promoted excessive ethanol drinking in ENT1(+/+) mice, but not in ENT1(-/-) mice. Our results indicate that A2AR-mediated CREB signaling in the DMS is a key determinant in enhancing the development of goal-directed ethanol drinking in mice.
Figures




References
-
- Arolfo MP, Yao L, Gordon AS, Diamond I, Janak PH. Ethanol operant self-administration in rats is regulated by adenosine A2 receptors. Alcohol Clin Exp Res. 2004;28:1308–1316. - PubMed
-
- Silva AJ, Simpson EM, Takahashi JS, Lipp HP, Nakanishi S, Wehner JM, Giese KP, Tully T, Abel T, Chapman PF, Fox K, Grant S, Itohara S, Lathe R, Mayford M, McNamara JO, Morris RJ, Picciotto M, Roder J, Shin HS, et al. Mutant mice and neuroscience: recommendations concerning genetic background. Neuron. 1997;19:755–759. - PubMed
-
- Carlezon WA, Jr, Thome J, Olson VG, Lane-Ladd SB, Brodkin ES, Hiroi N, Duman RS, Neve RL, Nestler EJ. Regulation of cocaine reward by CREB. Science. 1998;282:2272–2275. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Medical
Molecular Biology Databases