

Minor modifications to the phosphate groups and the C3' acyl chain length of lipid A in two Bordetella pertussis strains, BP338 and 18-323, independently affect Toll-like receptor 4 protein activation
- PMID: 23467413
- PMCID: PMC3636864
- DOI: 10.1074/jbc.M112.434365
Minor modifications to the phosphate groups and the C3' acyl chain length of lipid A in two Bordetella pertussis strains, BP338 and 18-323, independently affect Toll-like receptor 4 protein activation
Abstract
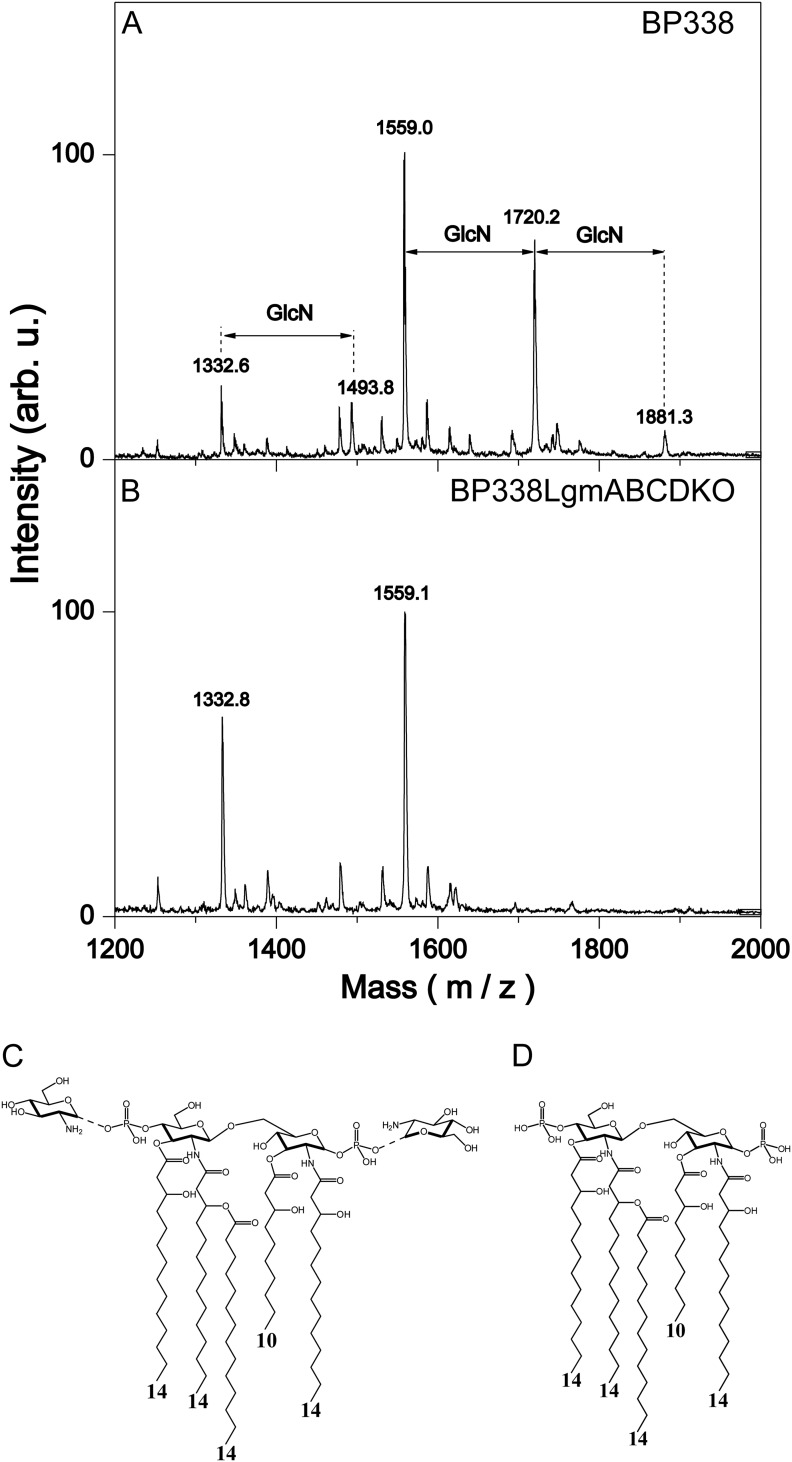
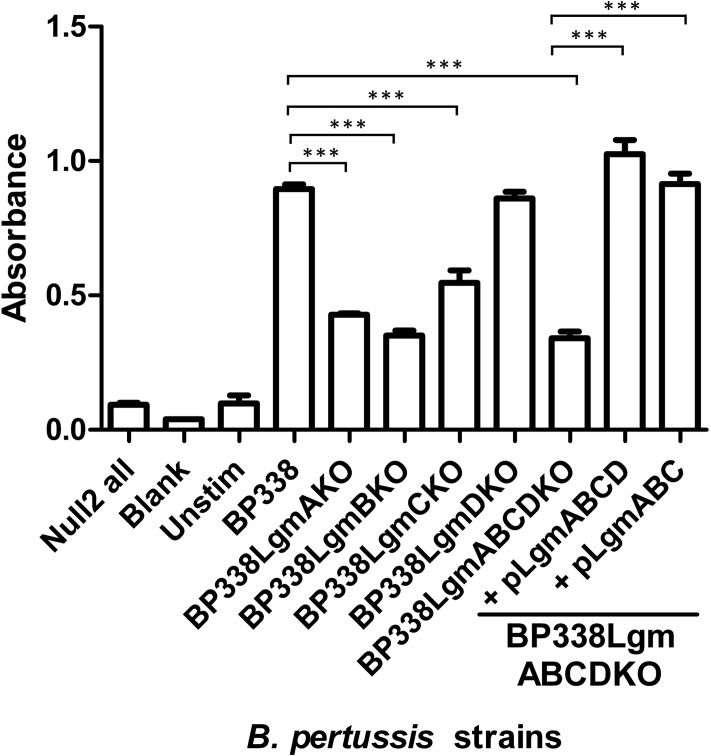
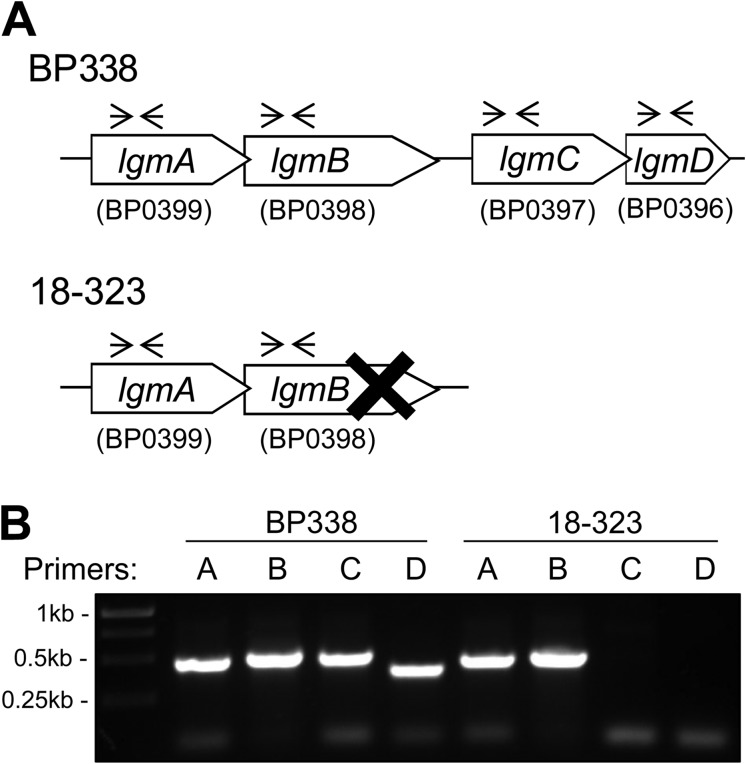
Lipopolysaccharides (LPS) of Bordetella pertussis are important modulators of the immune system. Interaction of the lipid A region of LPS with the Toll-like receptor 4 (TLR4) complex causes dimerization of TLR4 and activation of downstream nuclear factor κB (NFκB), which can lead to inflammation. We have previously shown that two strains of B. pertussis, BP338 (a Tohama I-derivative) and 18-323, display two differences in lipid A structure. 1) BP338 can modify the 1- and 4'-phosphates by the addition of glucosamine (GlcN), whereas 18-323 cannot, and 2) the C3' acyl chain in BP338 is 14 carbons long, but only 10 or 12 carbons long in 18-323. In addition, BP338 lipid A can activate TLR4 to a greater extent than 18-323 lipid A. Here we set out to determine the genetic reasons for the differences in these lipid A structures and the contribution of each structural difference to the ability of lipid A to activate TLR4. We show that three genes of the lipid A GlcN modification (Lgm) locus, lgmA, lgmB, and lgmC (previously locus tags BP0399-BP0397), are required for GlcN modification and a single amino acid difference in LpxA is responsible for the difference in C3' acyl chain length. Furthermore, by introducing lipid A-modifying genes into 18-323 to generate isogenic strains with varying penta-acyl lipid A structures, we determined that both modifications increase TLR4 activation, although the GlcN modification plays a dominant role. These results shed light on how TLR4 may interact with penta-acyl lipid A species.
Figures
References
-
- Trent M. S., Stead C. M., Tran A. X., Hankins J. V. (2006) Diversity of endotoxin and its impact on pathogenesis. J. Endotoxin Res. 12, 205–223 - PubMed
-
- Caroff M., Brisson J., Martin A., Karibian D. (2000) Structure of the Bordetella pertussis 1414 endotoxin. FEBS Lett. 477, 8–14 - PubMed
-
- Caroff M., Karibian D. (2003) Structure of bacterial lipopolysaccharides. Carbohydr. Res. 338, 2431–2447 - PubMed
-
- Trent M. S., Ribeiro A. A., Lin S., Cotter R. J., Raetz C. R. (2001) An inner membrane enzyme in Salmonella and Escherichia coli that transfers 4-amino-4-deoxy-l-arabinose to lipid A: induction on polymyxin-resistant mutants and role of a novel lipid-linked donor. J. Biol. Chem. 276, 43122–43131 - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Miscellaneous
