

The amidation step of diphthamide biosynthesis in yeast requires DPH6, a gene identified through mining the DPH1-DPH5 interaction network
- PMID: 23468660
- PMCID: PMC3585130
- DOI: 10.1371/journal.pgen.1003334
The amidation step of diphthamide biosynthesis in yeast requires DPH6, a gene identified through mining the DPH1-DPH5 interaction network
Abstract
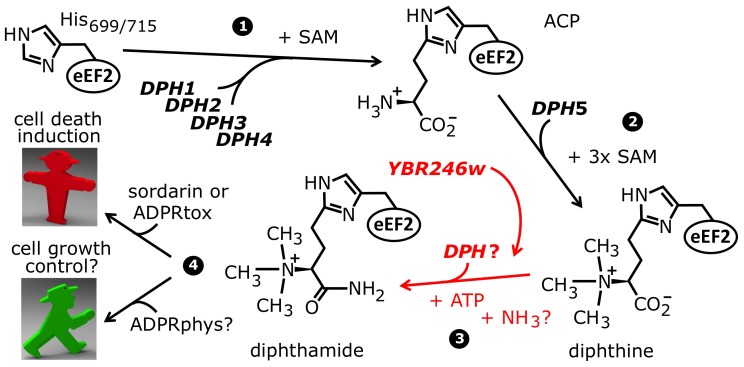
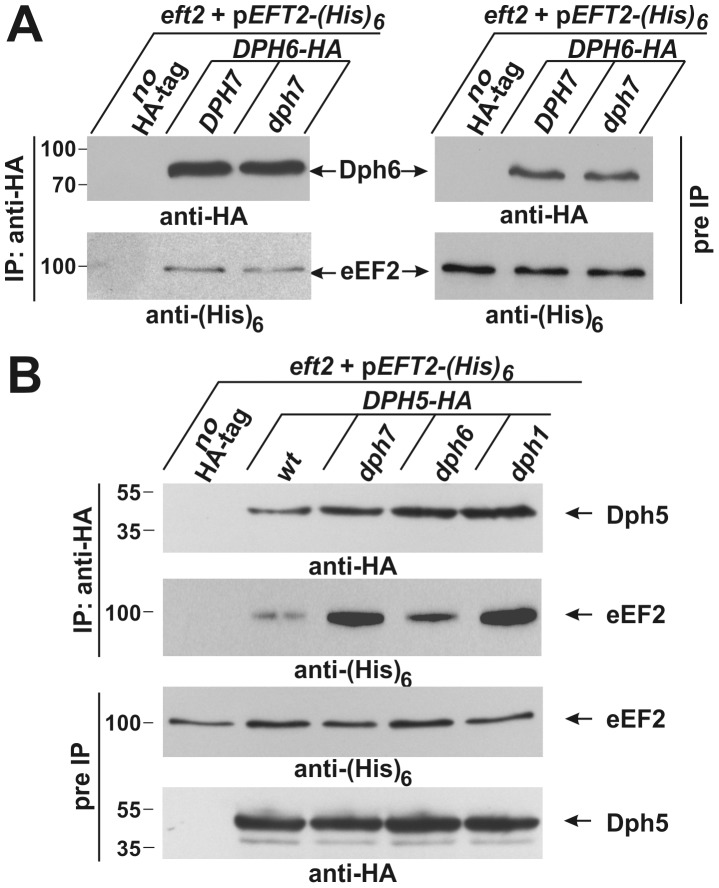
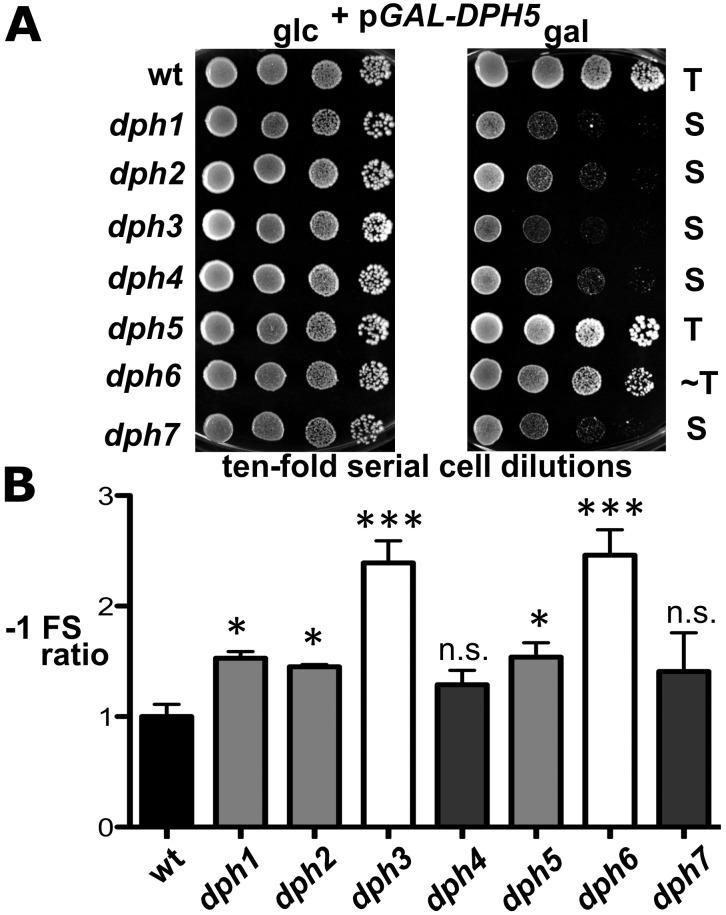
Diphthamide is a highly modified histidine residue in eukaryal translation elongation factor 2 (eEF2) that is the target for irreversible ADP ribosylation by diphtheria toxin (DT). In Saccharomyces cerevisiae, the initial steps of diphthamide biosynthesis are well characterized and require the DPH1-DPH5 genes. However, the last pathway step-amidation of the intermediate diphthine to diphthamide-is ill-defined. Here we mine the genetic interaction landscapes of DPH1-DPH5 to identify a candidate gene for the elusive amidase (YLR143w/DPH6) and confirm involvement of a second gene (YBR246w/DPH7) in the amidation step. Like dph1-dph5, dph6 and dph7 mutants maintain eEF2 forms that evade inhibition by DT and sordarin, a diphthamide-dependent antifungal. Moreover, mass spectrometry shows that dph6 and dph7 mutants specifically accumulate diphthine-modified eEF2, demonstrating failure to complete the final amidation step. Consistent with an expected requirement for ATP in diphthine amidation, Dph6 contains an essential adenine nucleotide hydrolase domain and binds to eEF2. Dph6 is therefore a candidate for the elusive amidase, while Dph7 apparently couples diphthine synthase (Dph5) to diphthine amidation. The latter conclusion is based on our observation that dph7 mutants show drastically upregulated interaction between Dph5 and eEF2, indicating that their association is kept in check by Dph7. Physiologically, completion of diphthamide synthesis is required for optimal translational accuracy and cell growth, as indicated by shared traits among the dph mutants including increased ribosomal -1 frameshifting and altered responses to translation inhibitors. Through identification of Dph6 and Dph7 as components required for the amidation step of the diphthamide pathway, our work paves the way for a detailed mechanistic understanding of diphthamide formation.
Conflict of interest statement
The authors have declared that no competing interests exist.
Figures





Comment in
-
Decoding the biosynthesis and function of diphthamide, an enigmatic modification of translation elongation factor 2 (EF2).Microb Cell. 2014 May 20;1(6):203-205. doi: 10.15698/mic2014.06.151. Microb Cell. 2014. PMID: 28357244 Free PMC article.
References
-
- Ahrne E, Muller M, Lisacek F (2010) Unrestricted identification of modified proteins using MS/MS. Proteomics 10: 671–686. - PubMed
-
- Seet BT, Dikic I, Zhou MM, Pawson T (2006) Reading protein modifications with interaction domains. Nat Rev Mol Cell Biol 7: 473–483. - PubMed
-
- Walsh CT, Garneau-Tsodikova S, Gatto GJJ (2005) Protein posttranslational modifications: the chemistry of proteome diversifications. Angew Chem Int Ed Engl 44: 7342–7372. - PubMed
-
- Greganova E, Altmann M, Butikofer P (2011) Unique modifications of translation elongation factors. FEBS J 278: 2613–2624. - PubMed
-
- Van Ness BG, Howard JB, Bodley JW (1980) ADP-ribosylation of elongation factor 2 by diphtheria toxin. NMR spectra and proposed structures of ribosyl-diphthamide and its hydrolysis products. J Biol Chem 255: 10710–10716. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
- 083524/Z/07/Z/WT_/Wellcome Trust/United Kingdom
- 097945/WT_/Wellcome Trust/United Kingdom
- BB/F019106/1/BB_/Biotechnology and Biological Sciences Research Council/United Kingdom
- BB/F0191629/1/BB_/Biotechnology and Biological Sciences Research Council/United Kingdom
- 1ZIAAI000929-09/PHS HHS/United States
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
Miscellaneous
