

Pregnancy-specific glycoproteins bind integrin αIIbβ3 and inhibit the platelet-fibrinogen interaction
- PMID: 23469002
- PMCID: PMC3585349
- DOI: 10.1371/journal.pone.0057491
Pregnancy-specific glycoproteins bind integrin αIIbβ3 and inhibit the platelet-fibrinogen interaction
Abstract
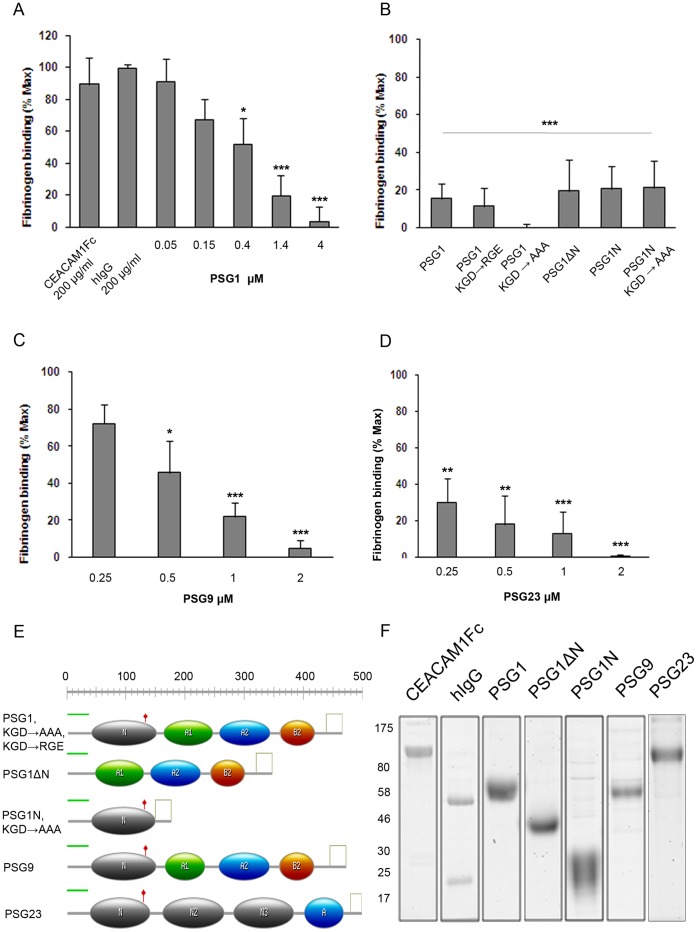
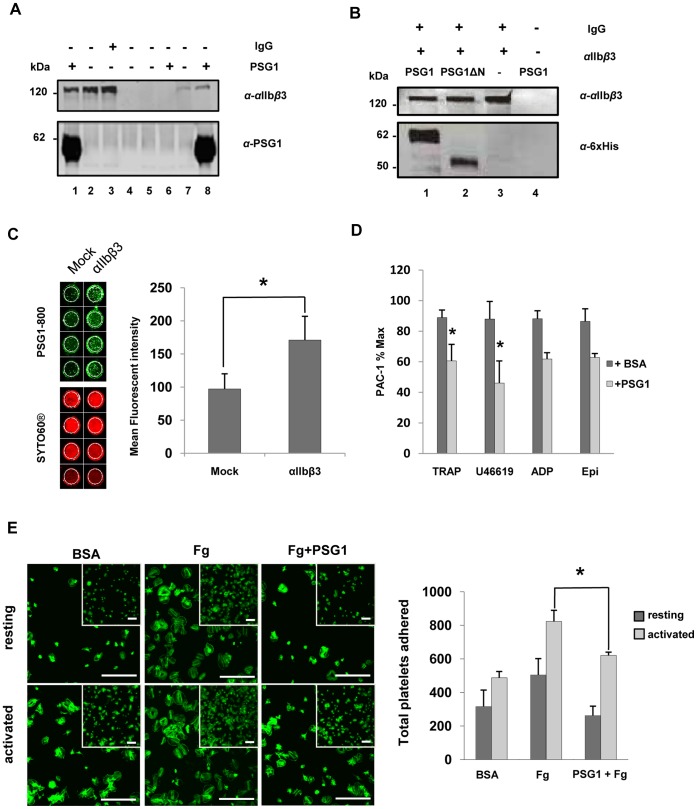
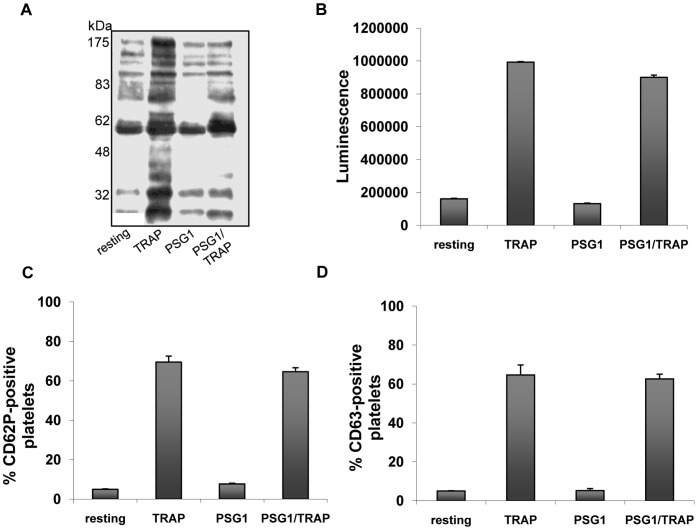
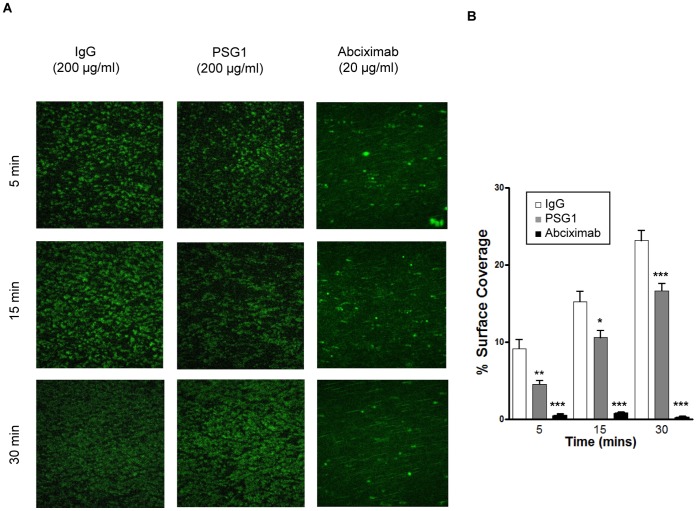
Pregnancy-specific glycoproteins (PSGs) are immunoglobulin superfamily members encoded by multigene families in rodents and primates. In human pregnancy, PSGs are secreted by the syncytiotrophoblast, a fetal tissue, and reach a concentration of up to 400 ug/ml in the maternal bloodstream at term. Human and mouse PSGs induce release of anti-inflammatory cytokines such as IL-10 and TGFβ1 from monocytes, macrophages, and other cell types, suggesting an immunoregulatory function. RGD tri-peptide motifs in the majority of human PSGs suggest that they may function like snake venom disintegrins, which bind integrins and inhibit interactions with ligands. We noted that human PSG1 has a KGD, rather than an RGD motif. The presence of a KGD in barbourin, a platelet integrin αIIbβ3 antagonist found in snake venom, suggested that PSG1 may be a selective αIIbβ3 ligand. Here we show that human PSG1 binds αIIbβ3 and inhibits the platelet - fibrinogen interaction. Unexpectedly, however, the KGD is not critical as multiple PSG1 domains independently bind and inhibit αIIbβ3 function. Human PSG9 and mouse Psg23 are also inhibitory suggesting conservation of this function across primate and rodent PSG families. Our results suggest that in species with haemochorial placentation, in which maternal blood is in direct contact with fetal trophoblast, the high expression level of PSGs reflects a requirement to antagonise abundant (3 mg/ml) fibrinogen in the maternal circulation, which may be necessary to prevent platelet aggregation and thrombosis in the prothrombotic maternal environment of pregnancy.
Conflict of interest statement
Figures
References
-
- Lin TM, Galbert SP, Kiefer D, Spellacy WN, Gall S (1974) Characterization of four human pregnancy-associated plasma proteins. Am J Obstet Gynecol 118: 223–236. - PubMed
-
- Towler CM, Horne CH, Jandial V, Campbell DM, MacGillivray I (1976) Plasma levels of pregnancy-specific beta1-glycoprotein in normal pregnancy. Br J Obstet Gynaecol. 83(10): 775–9. - PubMed
-
- Zhou GQ, Baranov V, Zimmermann W, Grunert F, Erhard B, et al. (1997) Highly specific monoclonal antibody demonstrates that pregnancy-specific glycoprotein (PSG) is limited to syncytiotrophoblast in human early and term placenta. Placenta 18(7): 491–501. - PubMed
-
- Teglund S, Zhou GQ, Hammarström S (1995) Characterization of cDNA encoding novel pregnancy-specific glycoprotein variants.Biochem Biophys Res Commun. 211(2): 656–664. - PubMed
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
