

Mass spectrometry of intact membrane protein complexes
- PMID: 23471109
- PMCID: PMC4058633
- DOI: 10.1038/nprot.2013.024
Mass spectrometry of intact membrane protein complexes
Abstract
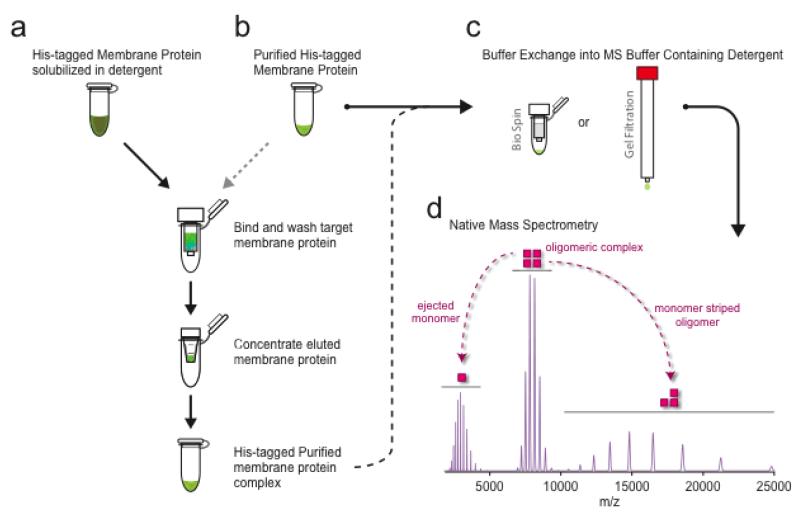
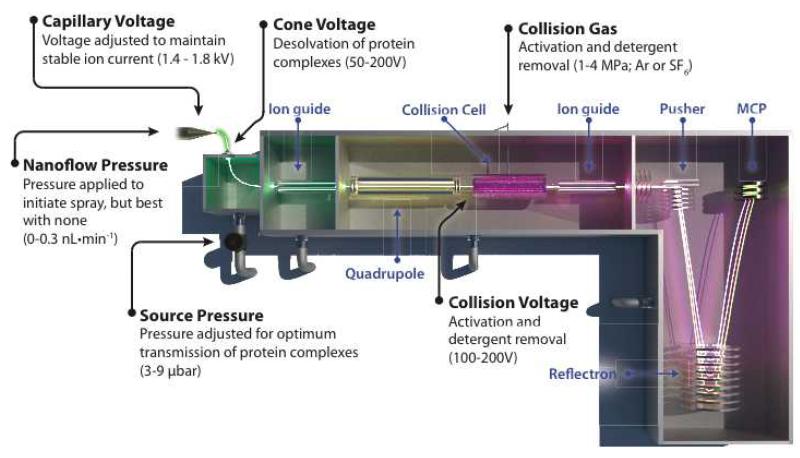
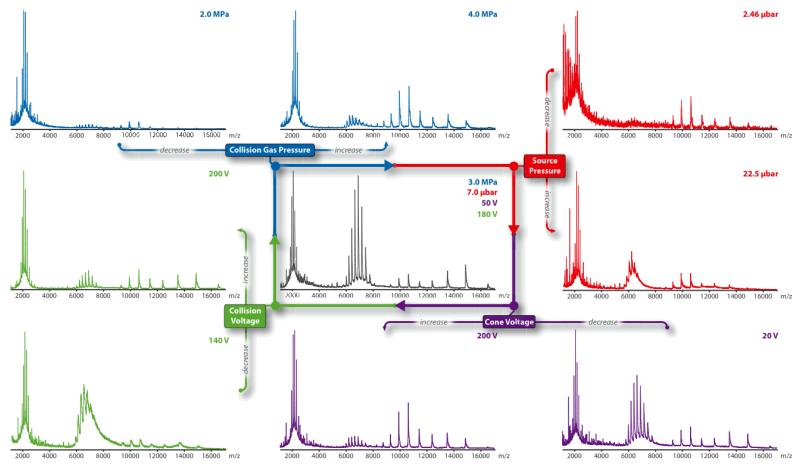
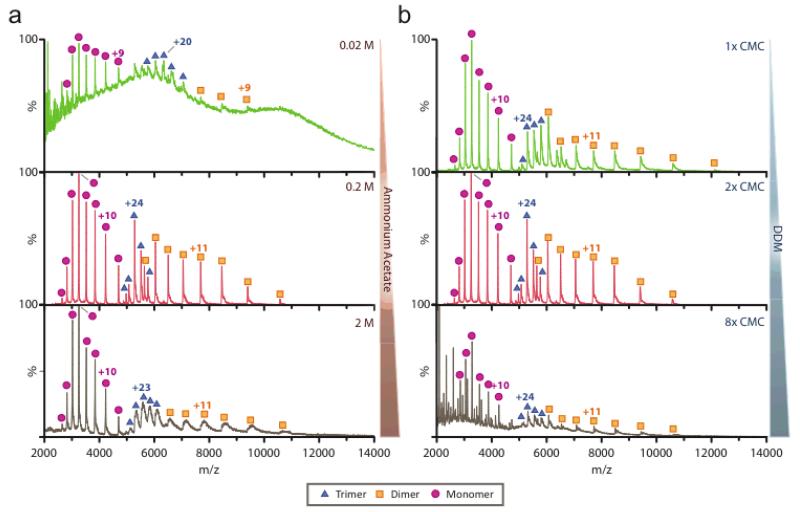
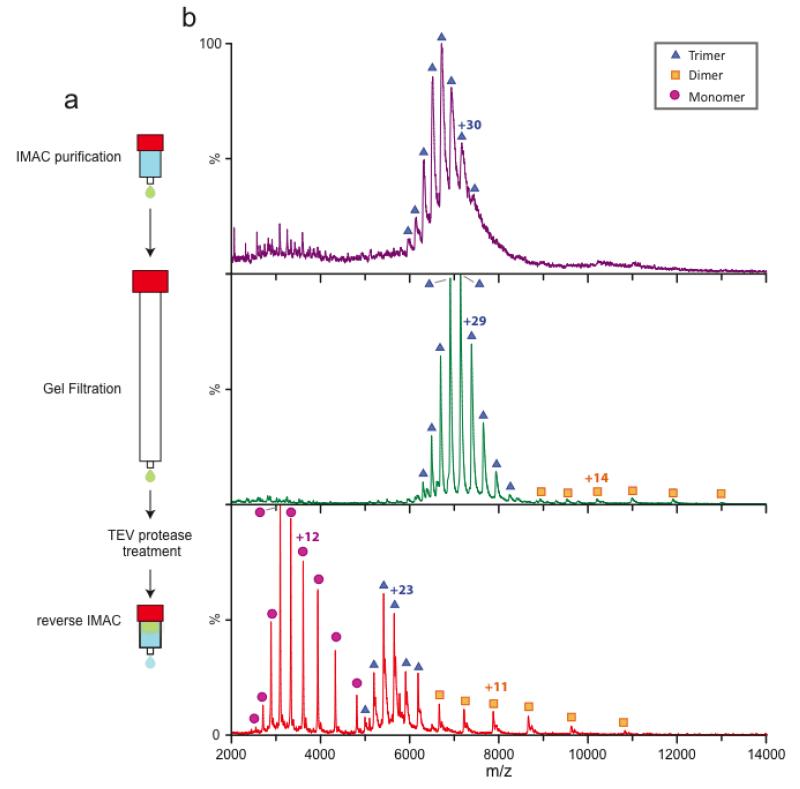
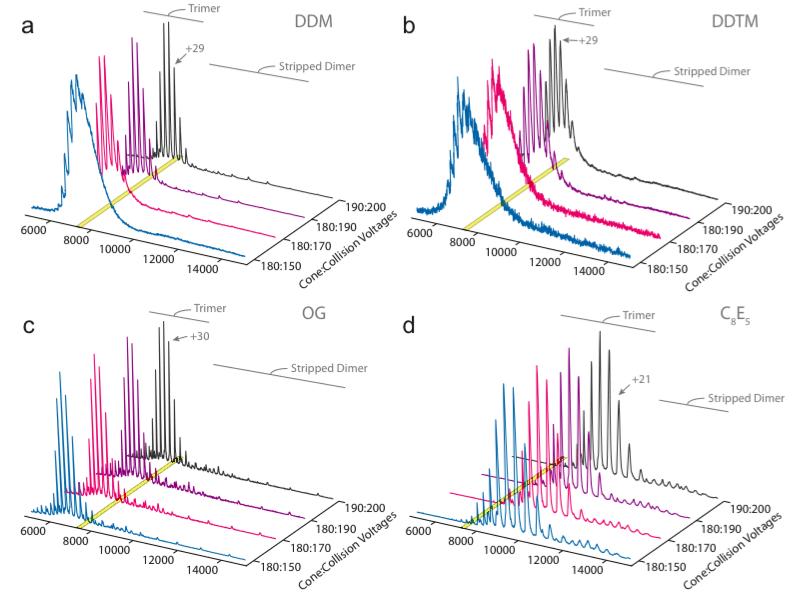
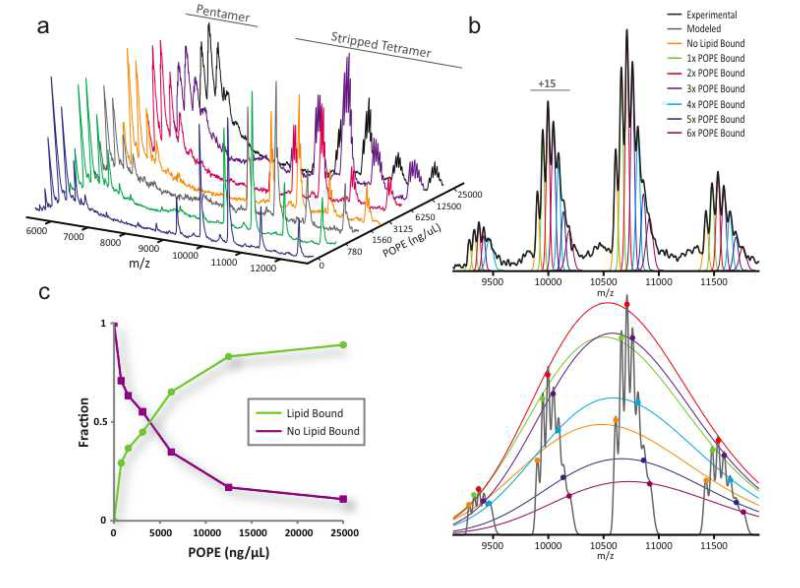
Mass spectrometry (MS) of intact soluble protein complexes has emerged as a powerful technique to study the stoichiometry, structure-function and dynamics of protein assemblies. Recent developments have extended this technique to the study of membrane protein complexes, where it has already revealed subunit stoichiometries and specific phospholipid interactions. Here we describe a protocol for MS of membrane protein complexes. The protocol begins with the preparation of the membrane protein complex, enabling not only the direct assessment of stoichiometry, delipidation and quality of the target complex but also the evaluation of the purification strategy. A detailed list of compatible nonionic detergents is included, along with a protocol for screening detergents to find an optimal one for MS, biochemical and structural studies. This protocol also covers the preparation of lipids for protein-lipid binding studies and includes detailed settings for a quadrupole time-of-flight (Q-TOF) mass spectrometer after the introduction of complexes from gold-coated nanoflow capillaries.
Figures
References
-
- Dorsam RT, Gutkind JS. G-protein-coupled receptors and cancer. Nat Rev Cancer. 2007;7:79–94. doi:10.1038/nrc2069. - PubMed
-
- Lappano R, Maggiolini M. G protein-coupled receptors: novel targets for drug discovery in cancer. Nat Rev Drug Discov. 2011;10:47–60. doi:10.1038/nrd3320. - PubMed
-
- Barrera NP, Di Bartolo N, Booth PJ, Robinson CV. Micelles protect membrane complexes from solution to vacuum. Science. 2008;321:243–246. doi:10.1126/science.1159292. - PubMed
-
- Barrera NP, Robinson CV. Advances in the mass spectrometry of membrane proteins: from individual proteins to intact complexes. Annu Rev Biochem. 2011;80:247–271. doi:10.1146/annurev-biochem-062309-093307. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
