

Molecular regulation of antibiotic biosynthesis in streptomyces
- PMID: 23471619
- PMCID: PMC3591988
- DOI: 10.1128/MMBR.00054-12
Molecular regulation of antibiotic biosynthesis in streptomyces
Abstract
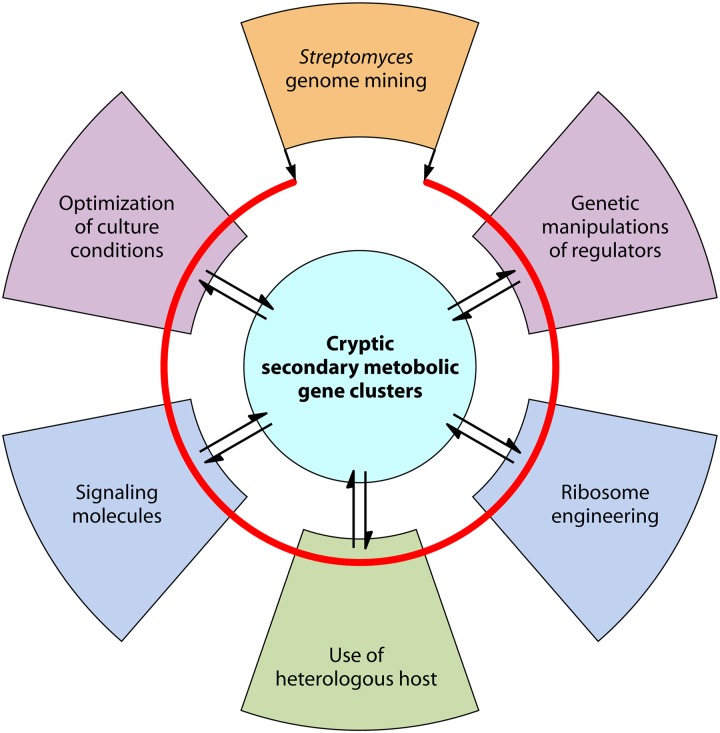
Streptomycetes are the most abundant source of antibiotics. Typically, each species produces several antibiotics, with the profile being species specific. Streptomyces coelicolor, the model species, produces at least five different antibiotics. We review the regulation of antibiotic biosynthesis in S. coelicolor and other, nonmodel streptomycetes in the light of recent studies. The biosynthesis of each antibiotic is specified by a large gene cluster, usually including regulatory genes (cluster-situated regulators [CSRs]). These are the main point of connection with a plethora of generally conserved regulatory systems that monitor the organism's physiology, developmental state, population density, and environment to determine the onset and level of production of each antibiotic. Some CSRs may also be sensitive to the levels of different kinds of ligands, including products of the pathway itself, products of other antibiotic pathways in the same organism, and specialized regulatory small molecules such as gamma-butyrolactones. These interactions can result in self-reinforcing feed-forward circuitry and complex cross talk between pathways. The physiological signals and regulatory mechanisms may be of practical importance for the activation of the many cryptic secondary metabolic gene cluster pathways revealed by recent sequencing of numerous Streptomyces genomes.
Figures
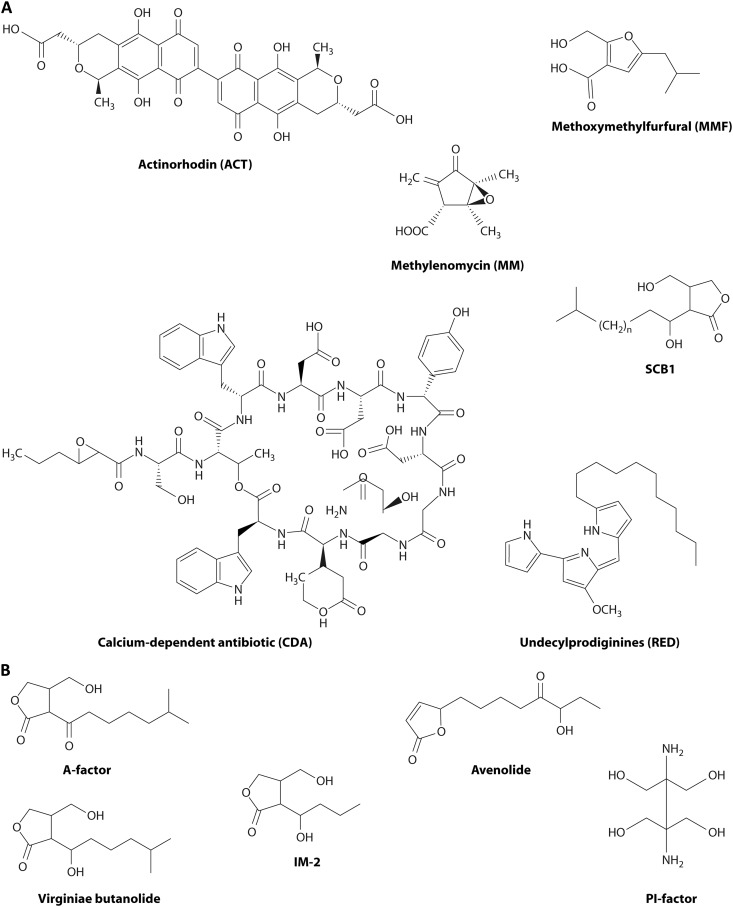
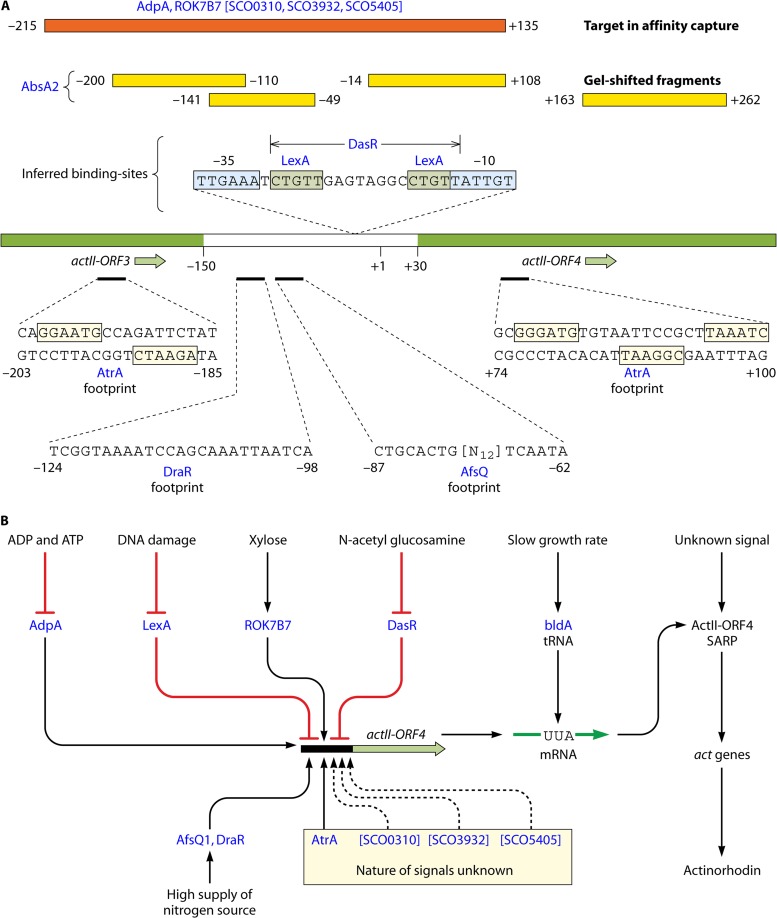
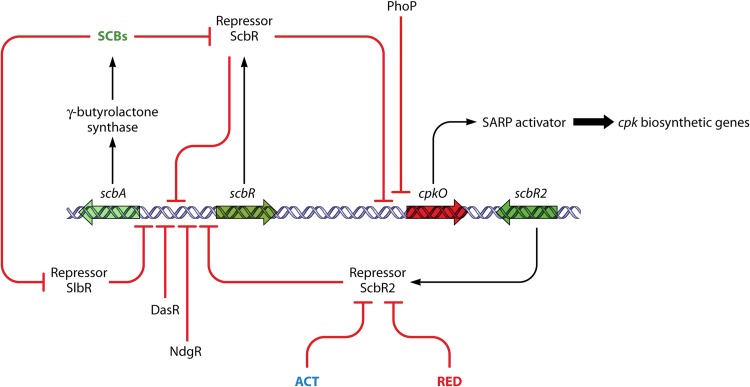
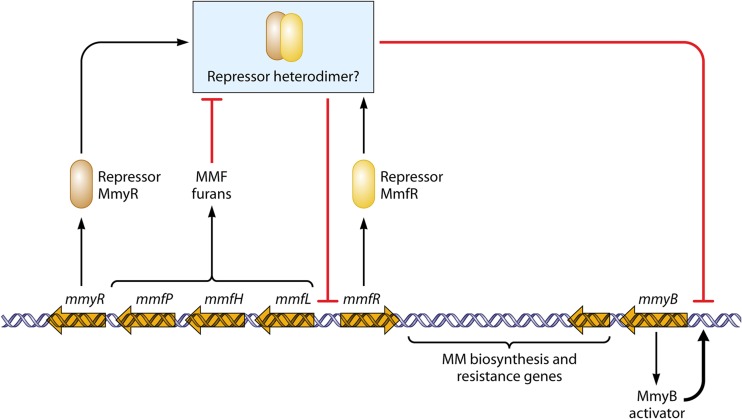
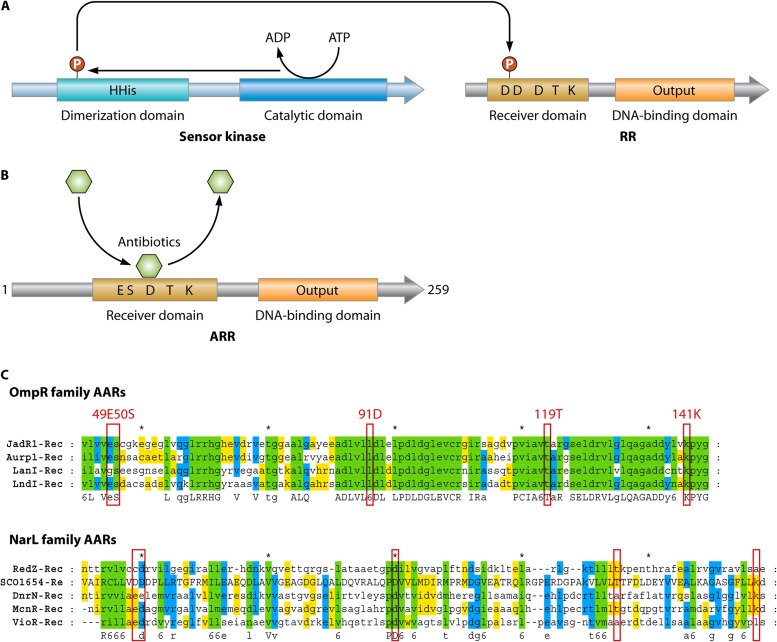
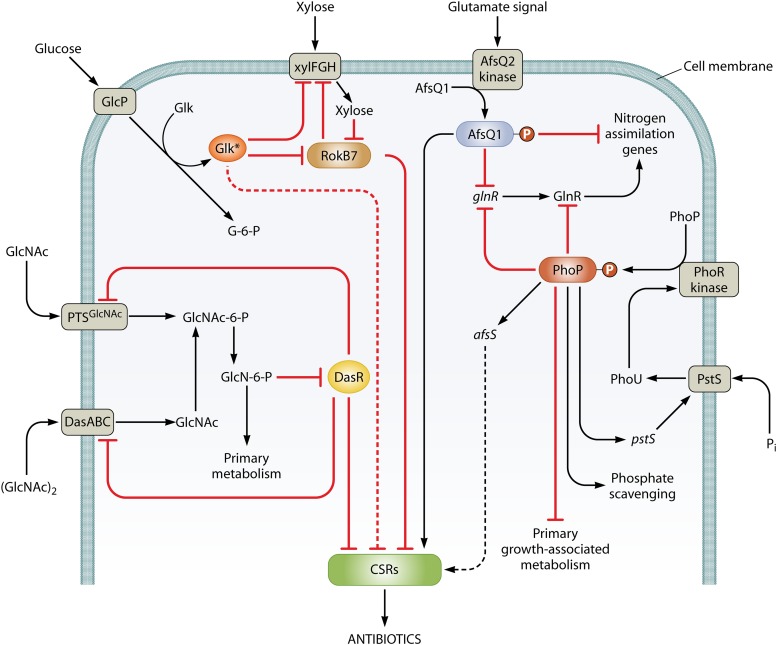
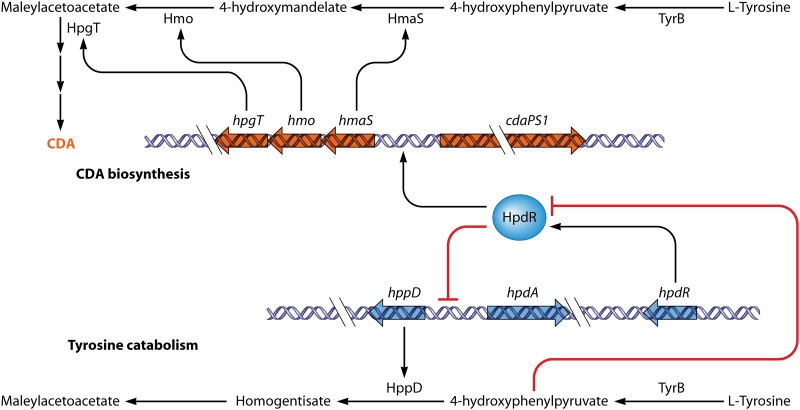
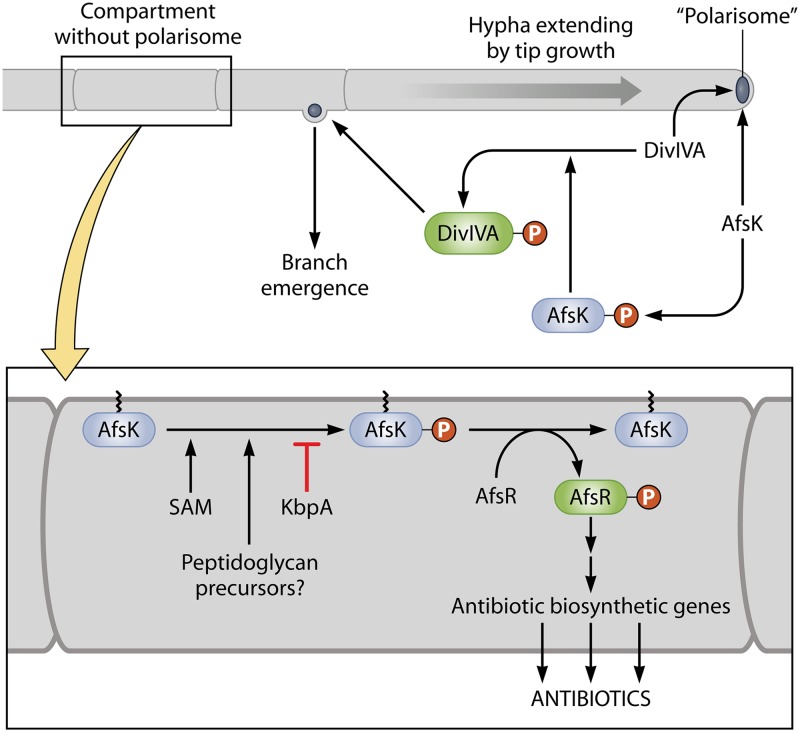
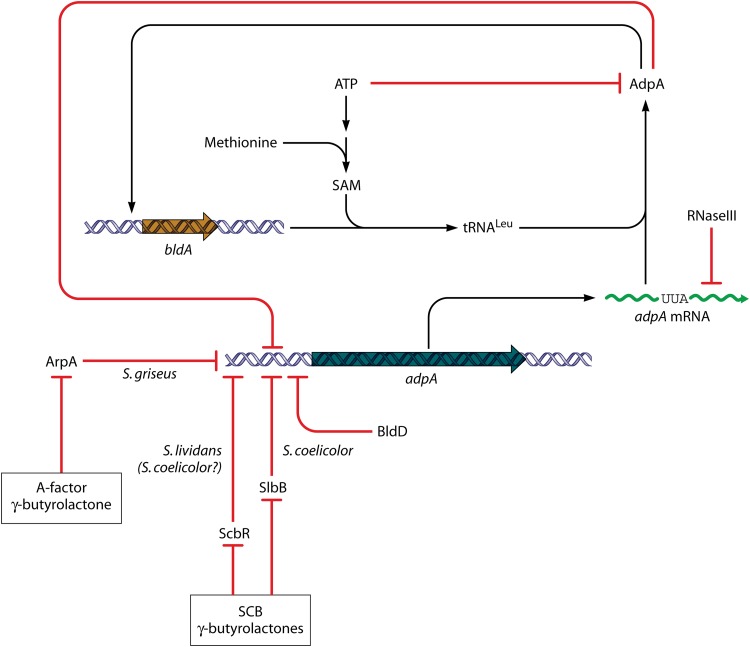
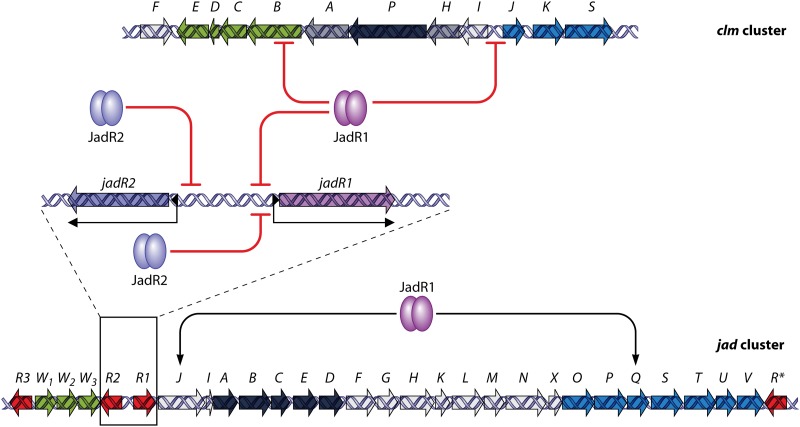
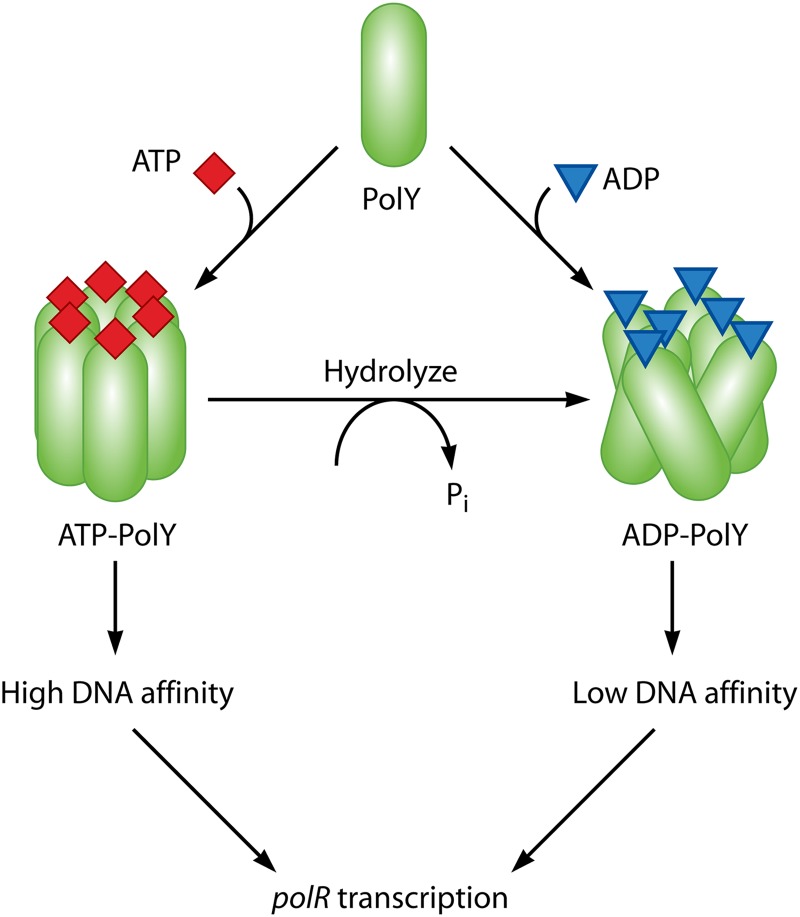
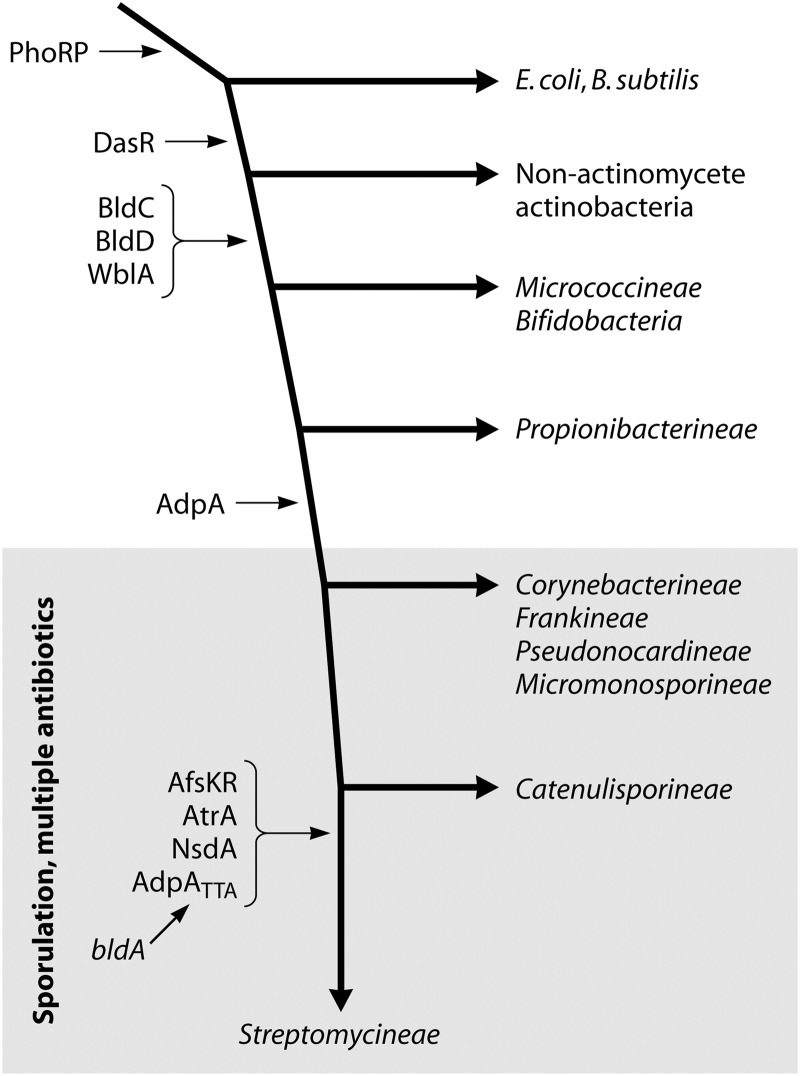





References
-
- Rudd BA. 1978. Genetics of pigmented secondary metabolites in Streptomyces coelicolor. University of East Anglia, Norwich, United Kingdom
-
- Rudd BA, Hopwood DA. 1979. Genetics of actinorhodin biosynthesis by Streptomyces coelicolor A3(2). J. Gen. Microbiol. 114: 35– 43 - PubMed
-
- Rudd BA, Hopwood DA. 1980. A pigmented mycelial antibiotic in Streptomyces coelicolor: control by a chromosomal gene cluster. J. Gen. Microbiol. 119: 333– 340 - PubMed
-
- Kirby R, Wright LF, Hopwood DA. 1975. Plasmid-determined antibiotic synthesis and resistance in Streptomyces coelicolor. Nature 254: 265– 267 - PubMed
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Other Literature Sources
Medical
Molecular Biology Databases
