

Chloroquine binding reveals flavin redox switch function of quinone reductase 2
- PMID: 23471972
- PMCID: PMC3630856
- DOI: 10.1074/jbc.M113.457002
Chloroquine binding reveals flavin redox switch function of quinone reductase 2
Abstract
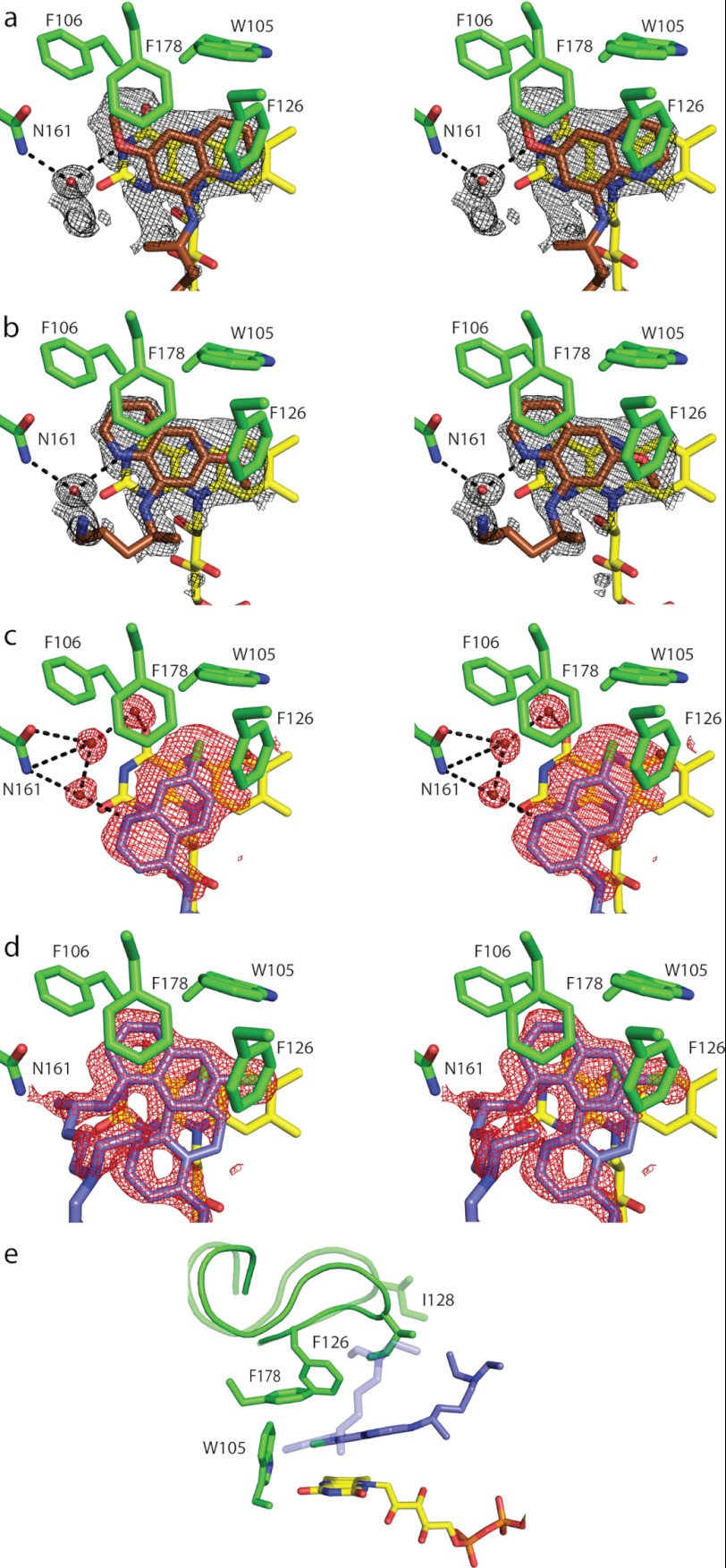
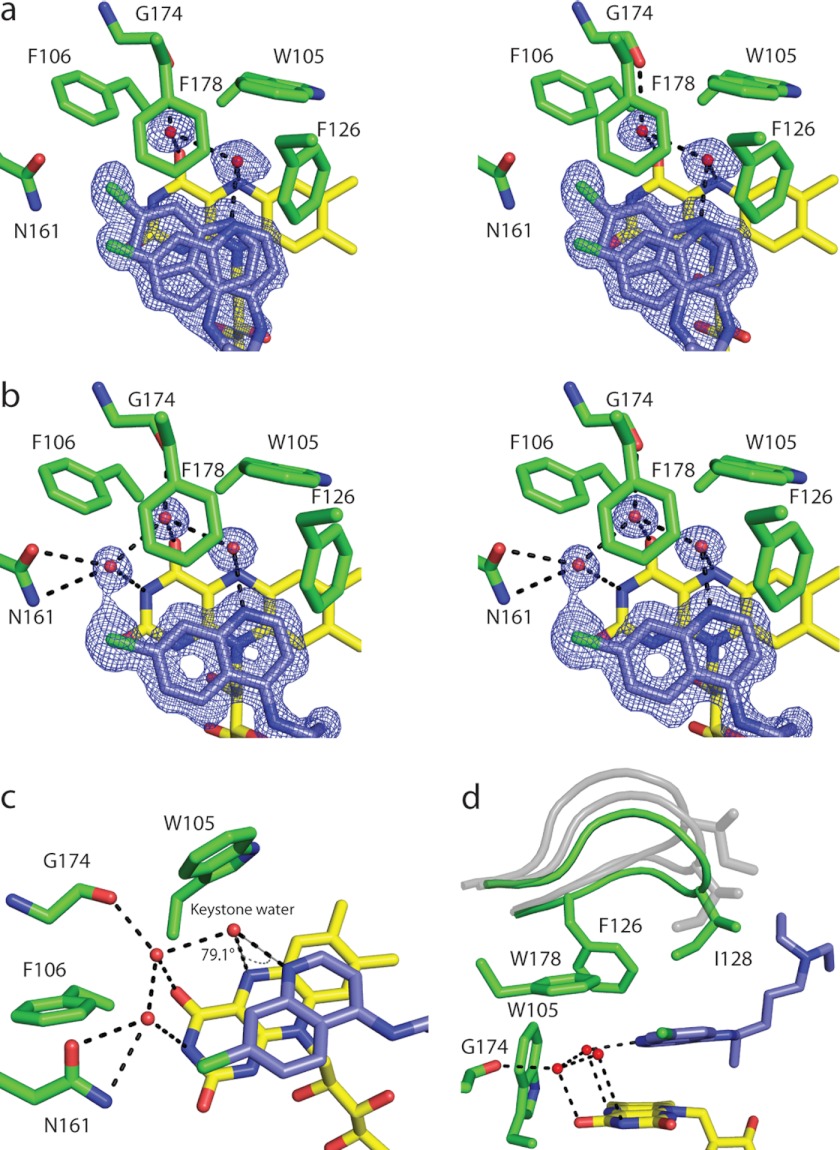
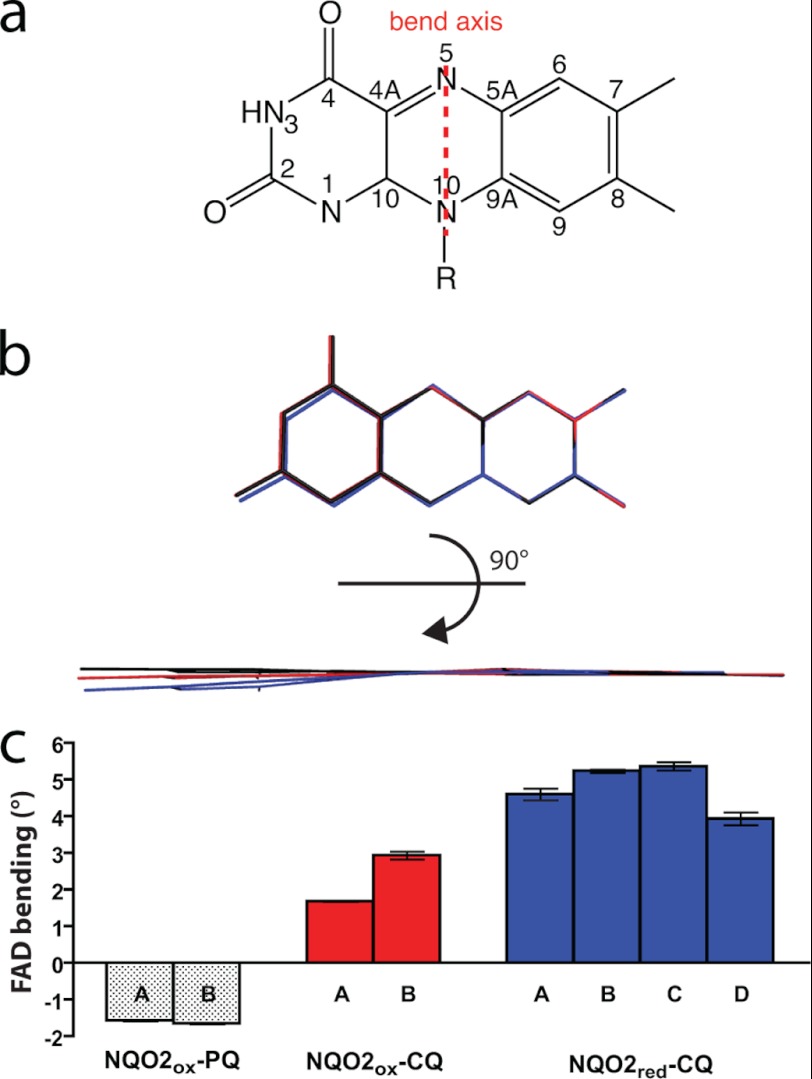
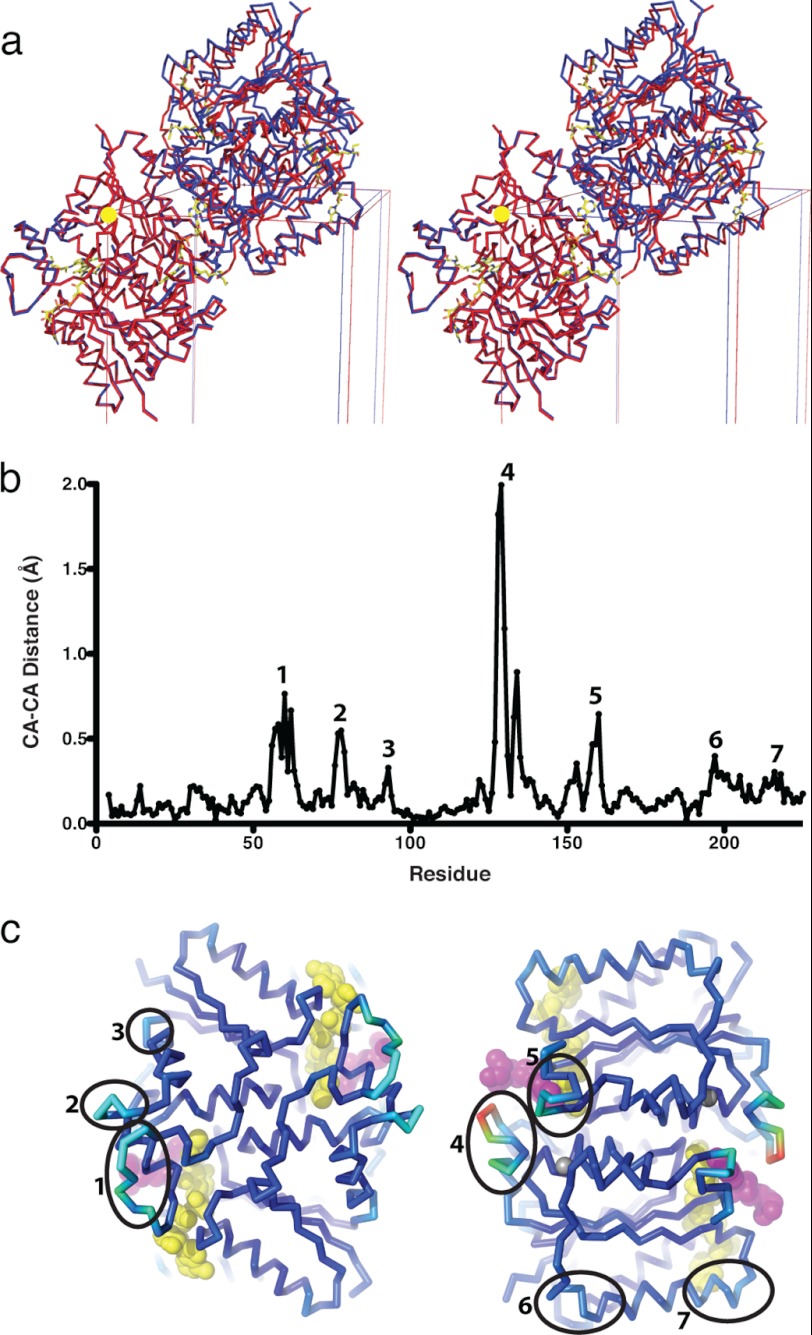
Quinone reductase 2 (NQO2) is an FAD-linked enzyme and the only known human target of two antimalarial drugs, primaquine (PQ) and chloroquine (CQ). The structural differences between oxidized and reduced NQO2 and the structural basis for inhibition by PQ and CQ were investigated by x-ray crystallography. Structures of oxidized NQO2 in complex with PQ and CQ were solved at 1.4 Å resolution. CQ binds preferentially to reduced NQO2, and upon reduction of NQO2-CQ crystals, the space group changed from P2(1)2(1)2(1) to P2(1), with 1-Å decreases in all three unit cell dimensions. The change in crystal packing originated in the negative charge and 4-5º bend in the reduced isoalloxazine ring of FAD, which resulted in a new mode of CQ binding and closure of a flexible loop (Phe(126)-Leu(136)) over the active site. This first structure of a reduced quinone reductase shows that reduction of the FAD cofactor and binding of a specific inhibitor lead to global changes in NQO2 structure and is consistent with a functional role for NQO2 as a flavin redox switch.
Figures
References
-
- Hill D. R., Baird J. K., Parise M. E., Lewis L. S., Ryan E. T., Magill A. J. (2006) Primaquine. Report from CDC expert meeting on malaria chemoprophylaxis I. Am. J. Trop. Med. Hyg. 75, 402–415 - PubMed
-
- Lalloo D. G., Shingadia D., Pasvol G., Chiodini P. L., Whitty C. J., Beeching N. J., Hill D. R., Warrell D. A., Bannister B. A., and HPA Advisory Committee on Malaria Prevention in UK Travellers (2007) UK malaria treatment guidelines. J. Infect. 54, 111–121 - PubMed
-
- Foley M., Tilley L. (1998) Quinoline antimalarials. Mechanisms of action and resistance and prospects for new agents. Pharmacol. Ther. 79, 55–87 - PubMed
-
- Fitch C. D. (2004) Ferriprotoporphyrin IX, phospholipids, and the antimalarial actions of quinoline drugs. Life Sci. 74, 1957–1972 - PubMed
-
- Vale N., Moreira R., Gomes P. (2009) Primaquine revisited six decades after its discovery. Eur. J. Med. Chem. 44, 937–953 - PubMed
Publication types
MeSH terms
Substances
Associated data
- Actions
- Actions
- Actions
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
