

The insecticidal neurotoxin Aps III is an atypical knottin peptide that potently blocks insect voltage-gated sodium channels
- PMID: 23473802
- PMCID: PMC3654253
- DOI: 10.1016/j.bcp.2013.02.030
The insecticidal neurotoxin Aps III is an atypical knottin peptide that potently blocks insect voltage-gated sodium channels
Abstract
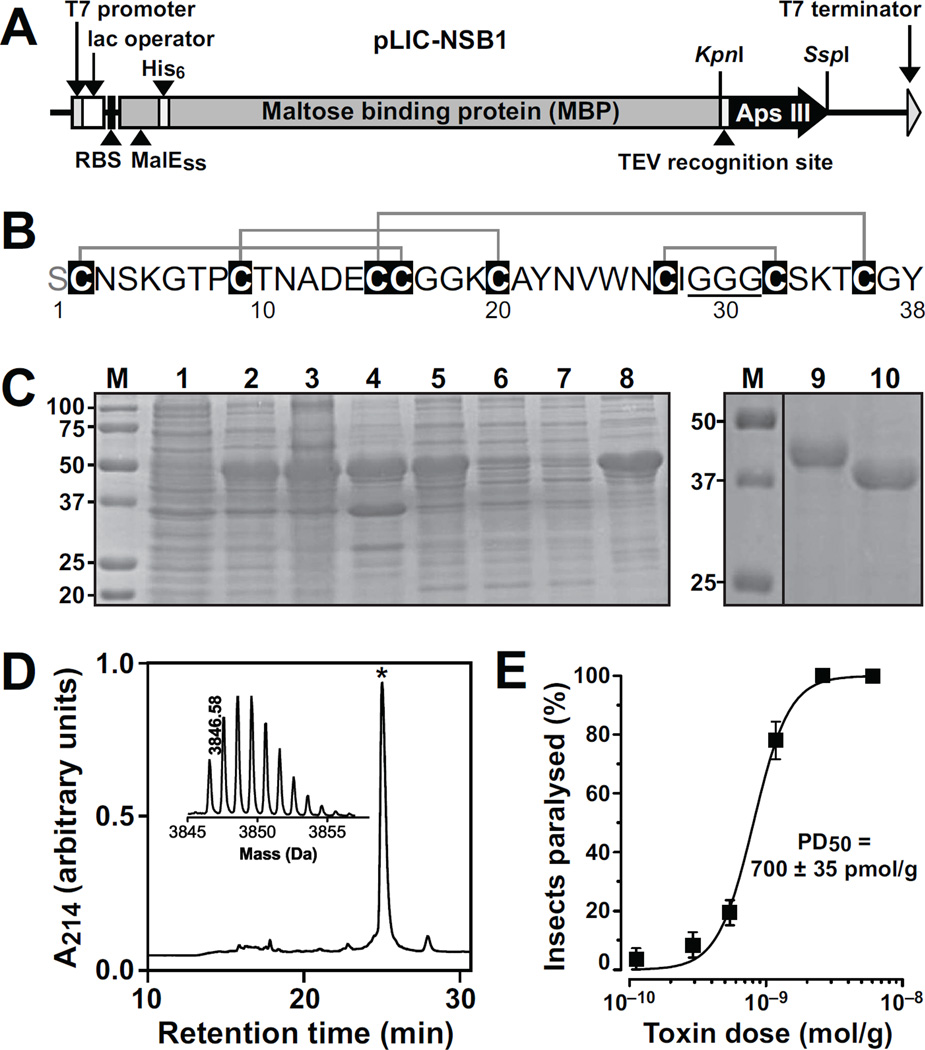
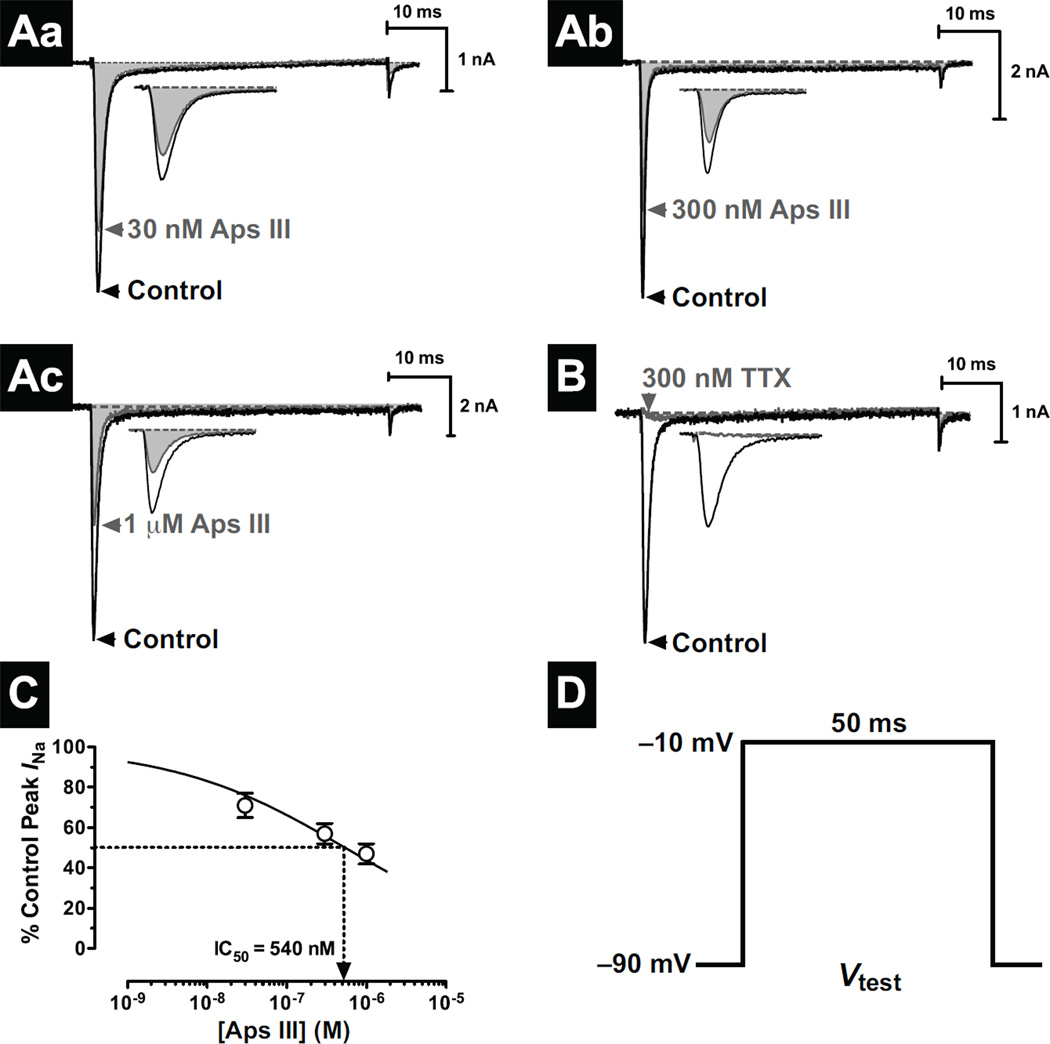
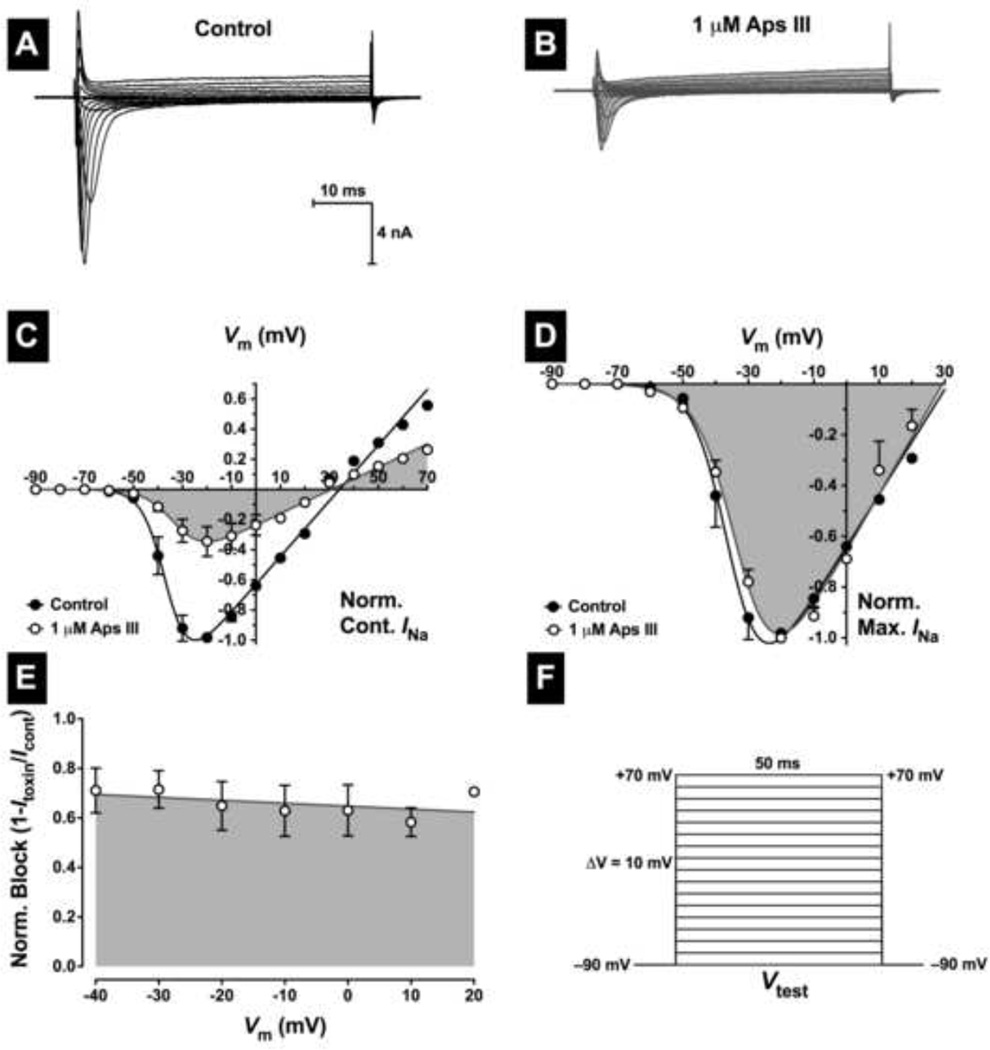
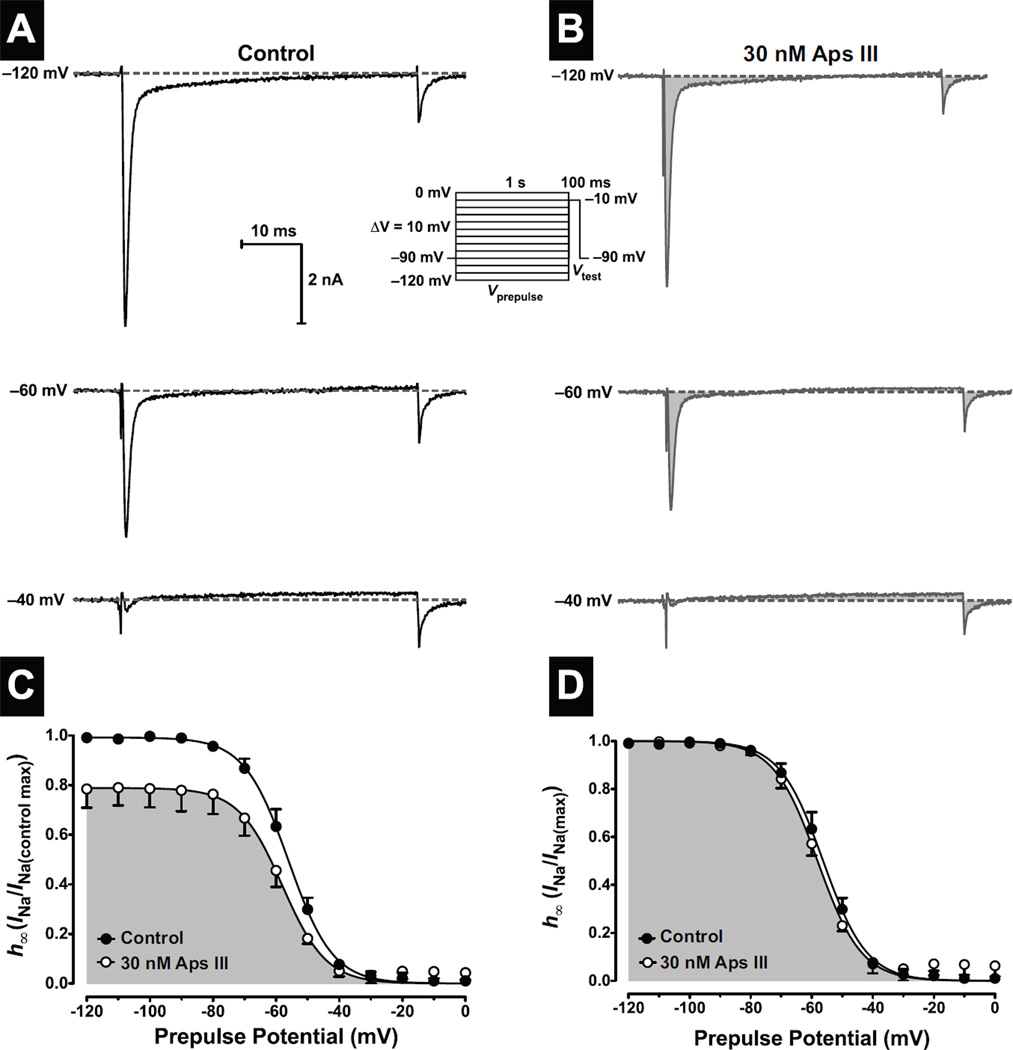
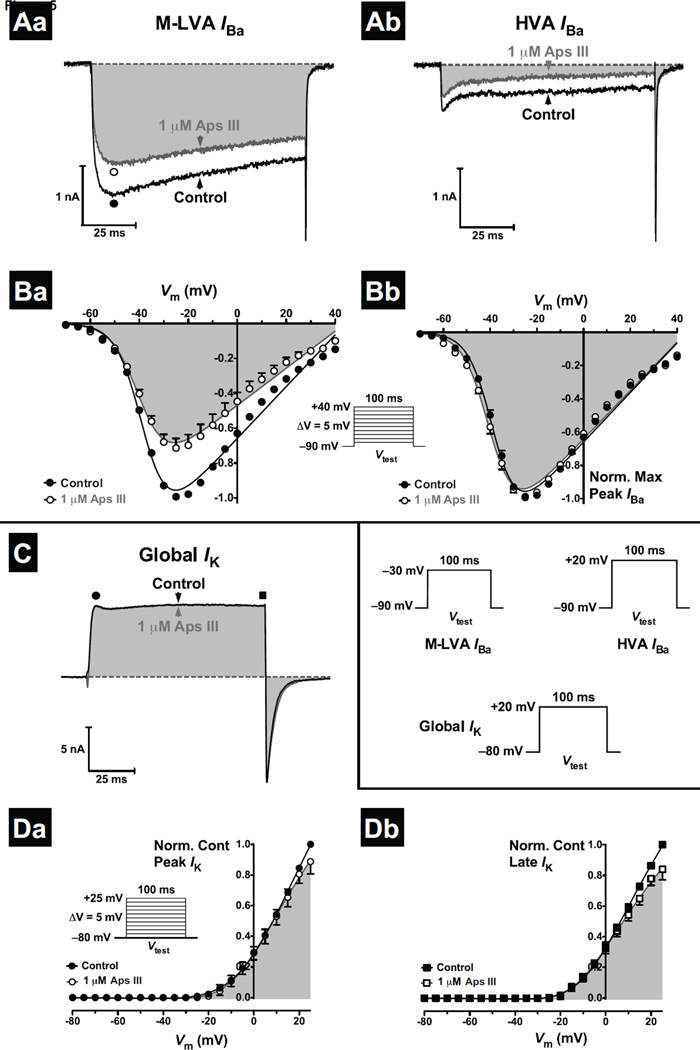
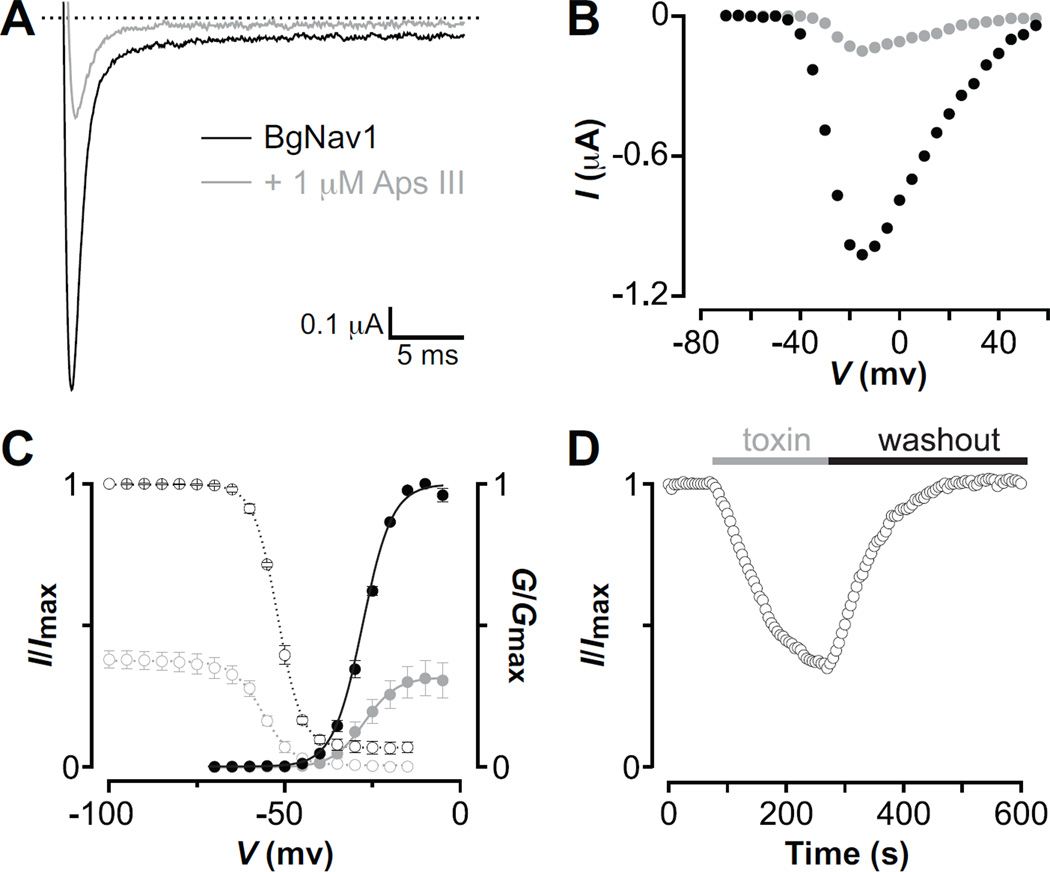
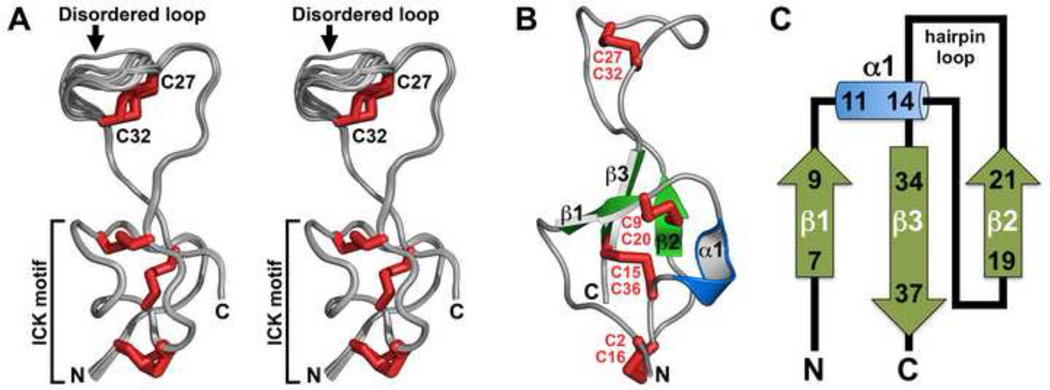
One of the most potent insecticidal venom peptides described to date is Aps III from the venom of the trapdoor spider Apomastus schlingeri. Aps III is highly neurotoxic to lepidopteran crop pests, making it a promising candidate for bioinsecticide development. However, its disulfide-connectivity, three-dimensional structure, and mode of action have not been determined. Here we show that recombinant Aps III (rAps III) is an atypical knottin peptide; three of the disulfide bridges form a classical inhibitor cystine knot motif while the fourth disulfide acts as a molecular staple that restricts the flexibility of an unusually large β hairpin loop that often houses the pharmacophore in this class of toxins. We demonstrate that the irreversible paralysis induced in insects by rAps III results from a potent block of insect voltage-gated sodium channels. Channel block by rAps III is voltage-independent insofar as it occurs without significant alteration in the voltage-dependence of channel activation or steady-state inactivation. Thus, rAps III appears to be a pore blocker that plugs the outer vestibule of insect voltage-gated sodium channels. This mechanism of action contrasts strikingly with virtually all other sodium channel modulators isolated from spider venoms that act as gating modifiers by interacting with one or more of the four voltage-sensing domains of the channel.
Copyright © 2013 Elsevier Inc. All rights reserved.
Conflict of interest statement
The authors declare that they have no conflicts of interest.
Figures
References
-
- Tedford HW, Sollod BL, Maggio F, King GF. Australian funnel-web spiders: master insecticide chemists. Toxicon. 2004;43:601–618. - PubMed
-
- World Health Organization. World Malaria Report. Geneva, Switzerland: WHO Press; 2009.
-
- Oerke EC. Crop losses to pests. J Agric Sci. 2006;144:31–43.
-
- Pimental D. Pesticides and pest control. In: Peshin R, Dhawan AK, editors. Integrated pest management: innovation-development process. Dordrecht: Springer Verlag; 2009. pp. 83–87.
-
- Tedford HW, Gilles N, Ménez A, Doering CJ, Zamponi GW, King GF. Scanning mutagenesis of ω-atracotoxin-Hv1a reveals a spatially restricted epitope that confers selective activity against invertebrate calcium channels. J Biol Chem. 2004;279:44133–44140. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
Miscellaneous
