

Review
doi: 10.1038/nrmicro2992.
Epub 2013 Mar 11.
A cornucopia of human polyomaviruses
Affiliations
- PMID: 23474680
- PMCID: PMC3928796
- DOI: 10.1038/nrmicro2992
Item in Clipboard
Review
A cornucopia of human polyomaviruses
Nat Rev Microbiol.
2013 Apr.
Abstract
During the past 6 years, focused virus hunting has led to the discovery of nine new human polyomaviruses, including Merkel cell polyomavirus, which has been linked to Merkel cell carcinoma, a lethal skin cell cancer. The discovery of so many new and highly divergent human polyomaviruses raises key questions regarding their evolution, tropism, latency, reactivation, immune evasion and contribution to disease. This Review describes the similarities and differences among the new human polyomaviruses and discusses how these viruses might interact with their human host.
Figures

a | The prototypical circular double-stranded DNA genome has three main regions: a non-coding control region containing the early and late promoters, their transcription start sites and the origin of replication; an early region encoding large T antigen (LT) and small T antigen (ST) and an alternatively spliced LT (LT′); and a late region encoding the viral coat proteins VP1, VP2, VP3 and VP4. The reading frames for VP2, VP3 and VP4 are identical, but translation starts at successive initiating AUG codons to generate the different proteins. VP4 has been confirmed only in the primate species simian virus 40 (SV40). Agnoprotein (Agno) is encoded by a late transcript from JC polyomavirus and BK polyomavirus, but has yet to be confirmed in the new human polyomaviruses. b | Electron micrograph of purified recombinant VP1 pentamers (from murine polyomavirus), which have a diameter of ~9 nm (scale bar represents 100 nm). c | Electron micrograph of purified murine polyomavirus virions, which have a diameter of ~50 nm (scale bar represents 200 nm).
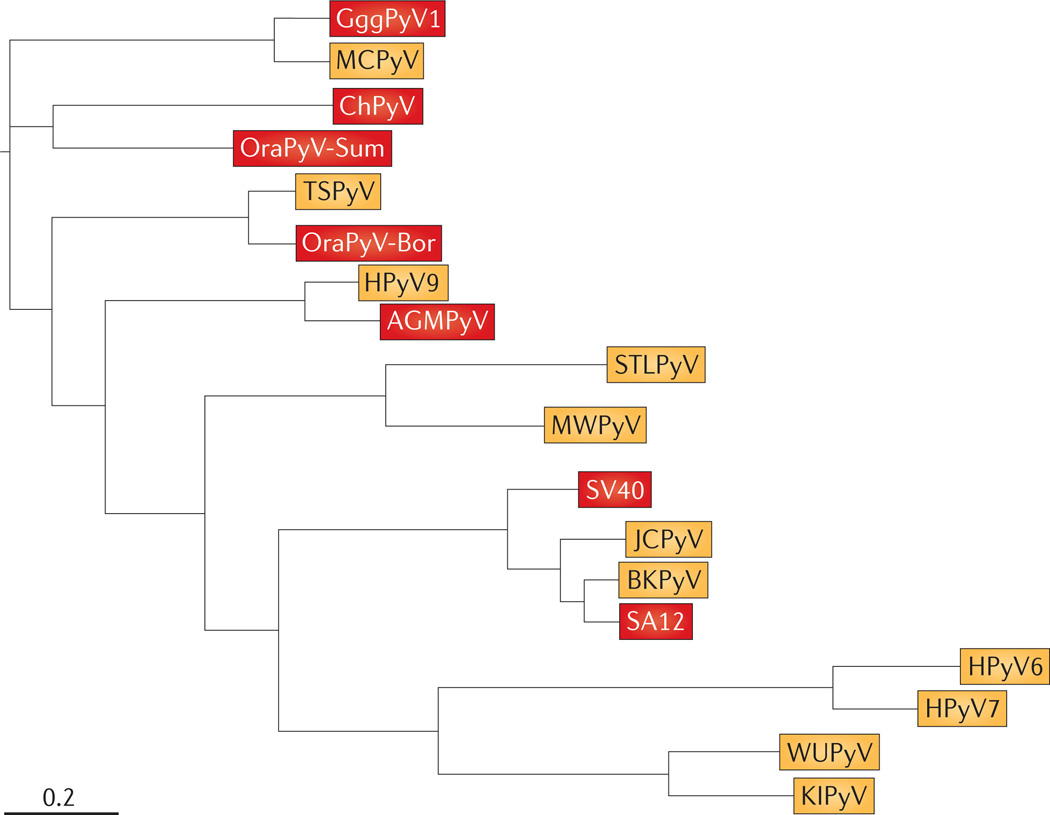
Viruses identified from human isolates are shown in yellow, and viruses identified from primate isolates are shown in red. This simplified tree was constructed by concatenating the amino acid sequences for the capsid protein VP1 and large T antigen from 19 separate isolates. Sequences were then aligned using MUSCLE 3.8.31 with default parameters. Alignment positions containing more than 90% gaps were filtered using filter_alignment.py from QIIME. The phylogenetic tree was then constructed using FastTree 2.1.4 with default parameters. Branch length is in substitutions per site. Note that Washington University polyomavirus (WUPyV), Karolinska Institute polyomavirus (KIPyV), human polyomavirus 6 (HPyV6) and HPyV7 all belong to the genus Wukipolyomavirus, and all other isolates shown belong to the genus Orthopolyomavirus. The RefSeq accession numbers for the human viruses are listed in TABLE 1. The primate viruses shown, and their GenBank or RefSeq accession numbers, are: Gorilla gorilla gorilla polyomavirus 1 (GggPyV1; HQ385752), chimpanzee polyomavirus (ChPyV; NC_014743), Sumatran orangutan polyomavirus (OraPyV-Sum; FN356901), Borneo OraPyV (OraPyV-Bor; NC_013439), African green monkey polyomavirus (AGMPyV; NC_004763), simian virus 40 (SV40; NC_001669) and simian agent 12 (SA12; NC_012122). Merkel cell polyomavirus (MCPyV) is also related to a number of chimpanzee isolates (not shown). Tree constructed by D. McDonald, University of Colorado at Boulder, USA. BKPyV, BK polyomavirus; JCPyV, JC polyomavirus; MWPyV, Malawi polyomavirus; STLPyV, St Louis polyomavirus; TSPyV, trichodysplasia spinulosa-associated polyomavirus.

a | The amino-terminal DnaJ (J) domain is shared between small and large T antigens. Large T antigen also contains a retinoblastoma-associated protein (RB)-binding LXCXE motif, a threonine–proline–proline–lysine (TPPK) motif, a nuclear-localization sequence (NLS), a DNA-binding domain (DBD), a helicase domain and a host range and adenovirus helper function (HR–AH) domain. Small T antigen contains an amino-terminal J domain followed by a unique region that is not shared with large T antigen and contains two zinc-fingers. b | Large T antigen binds to many cellular proteins. The J domain recruits heat shock cognate 71 kDa protein (HSC70) homologues. The region between the J domain and LXCXE motif has been shown to bind independently to cullin 7 (CUL7), BUB1 and insulin receptor substrate 1 (IRS1). Merkel cell polyomavirus (MCPyV) large T antigen binds to the endosomal protein VPS39. The LXCXE domain binds to retinoblastoma-associated protein (RB) and related proteins (p107 and p130). Phosphorylation of the threonine residue in the TPPK motif is required for large T antigen-mediated viral DNA replication. The nuclear localization signal (NLS) binds specifically to KPNA family importin homologues. The DBD and helicase domains are required for viral replication and recruit cellular DNA replication factors (DNA polymerase-α catalytic subunit (POLA), the replication protein A complex (RPA) and the DNA primase complex (PRIM) for DBD; and EP300, CREBBP, p53 and DNA topoisomerase 1 (TOP1) for the helicase domain) to the replicating simian virus 40 (SV40) genome. The carboxyl termini of SV40, JC polyomavirus (JCPyV) and BK polyomavirus (BKPyV) large T antigens each contain a threonine residue (threonine 701 in SV40, threonine 691 in BKPyV and threonine 684 in JCPyV) that, when phosphorylated, competes with phosphorylated G1–S-specific cyclin E1 and MYC for binding to the cullin RING ligase substrate adaptor FBXW7. FAM111A has recently been implicated in the SV40 host range restriction and adenovirus helper function. c | Small T antigen binds specifically to the A and C subunits of protein phosphatase 2A (PP2A). d | Polyomavirus genome replication. Replication begins when the large T antigen DBD binds to the origin of replication. Two hexamers of large T antigen form in a head-to-head orientation at the origin. The helicase domains of the large T antigen hexamers initiate unwinding of viral DNA followed by bidirectional replication.

Host glycans with different branching patterns are linked to lactosylceramide (Lac–Cer), which provides anchorage in the cell membrane. Although the affinity constants for a single glycan–VP1 interaction are low, the avidity of multiple interactions on the viral surface provides a stable attachment. Subtle modifications of the glycan are used by different viruses, as indicated, and several glycans have been co-crystallized in complex with the specific VP1 pentamer (see Functional Glycomics Gateway page for VP1 ). Glycan GT1b has been suggested as one glycan receptor for Merkel cell polyomavirus (MCPyV) on the basis of in vitro binding studies and X-ray crystallography,. It is also possible that receptor recognition changes for a particular virus, as demonstrated by the glycan preferences of ‘large plaque’ and ‘small plaque’ murine polyomavirus (MPyV).

a | In Merkel cell carcinoma, the Merkel cell polyomavirus (MCPyV) DNA undergoes mutations, leaving small T antigen and the J domain and LXCXE motif of large T antigen intact, but typically deleting the DNA-binding domain and helicase domain of large T antigen. b | An MCPyV genome (black) undergoes random integration into the host cell DNA (blue). If wild-type large T antigen is present in the cell, then repeated rounds of bidirectional viral DNA replication result in numerous copies of viral DNA, appearing as onion skinning, within the host chromosome. c | If a wild-type genome is integrated into the host and then obtains a mutation that inactivates large T antigen, the viral genome cannot be replicated bidirectionally and instead undergoes amplification in a manner analogous to amplification of cellular oncogenes such as MYC. d | An alternative model to explain the high copy number of integrated viral DNA is that rolling-circle amplification of an episomal MCPyV genome containing a mutation in the large T antigen gene results in a concatemer of mutant viral genomes. This concatemer then becomes randomly integrated into the host cell chromosome, resulting in a Merkel cell carcinoma containing a high copy number of viral genomes per cell.
References
-
- Groenewoud MJ, et al. Characterization of novel polyomaviruses from Bornean and Sumatran orangutans. J. Gen. Virol. 2010;91:653–658. - PubMed
Publication types
MeSH terms
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Medical
Miscellaneous
