

Aberrant Mer receptor tyrosine kinase expression contributes to leukemogenesis in acute myeloid leukemia
- PMID: 23474756
- PMCID: PMC3898106
- DOI: 10.1038/onc.2013.40
Aberrant Mer receptor tyrosine kinase expression contributes to leukemogenesis in acute myeloid leukemia
Erratum in
-
Aberrant Mer receptor tyrosine kinase expression contributes to leukemogenesis in acute myeloid leukemia.Oncogene. 2016 Dec 1;35(48):6270. doi: 10.1038/onc.2016.295. Epub 2016 Sep 5. Oncogene. 2016. PMID: 27593932 Free PMC article. No abstract available.
Abstract
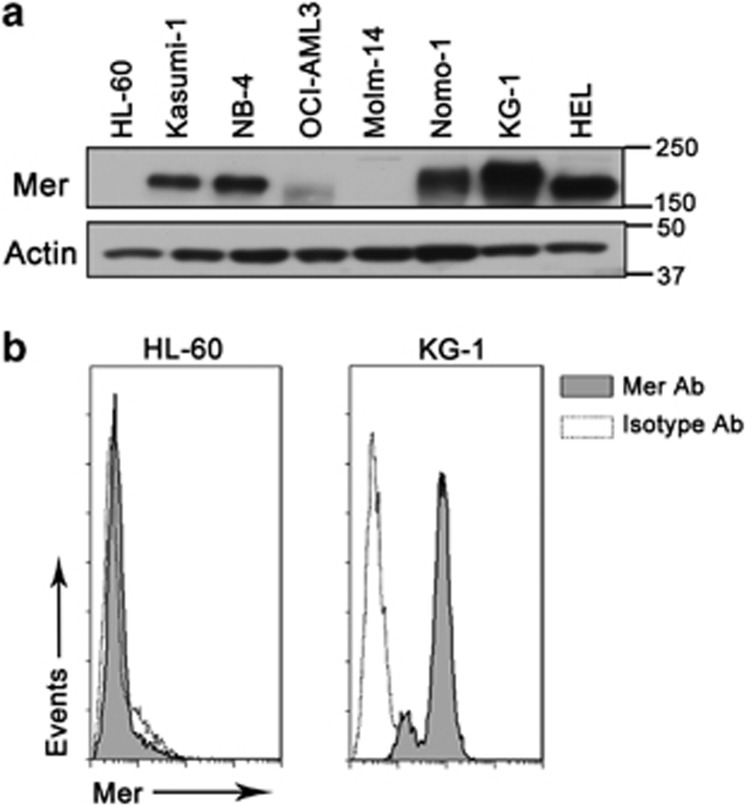
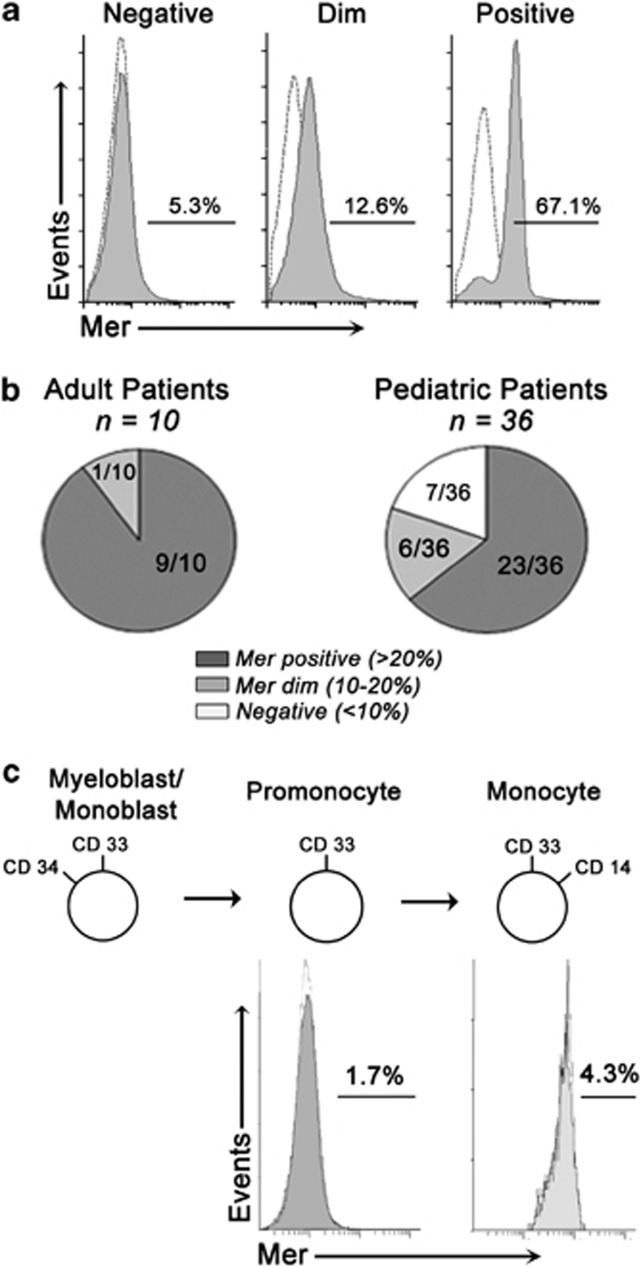
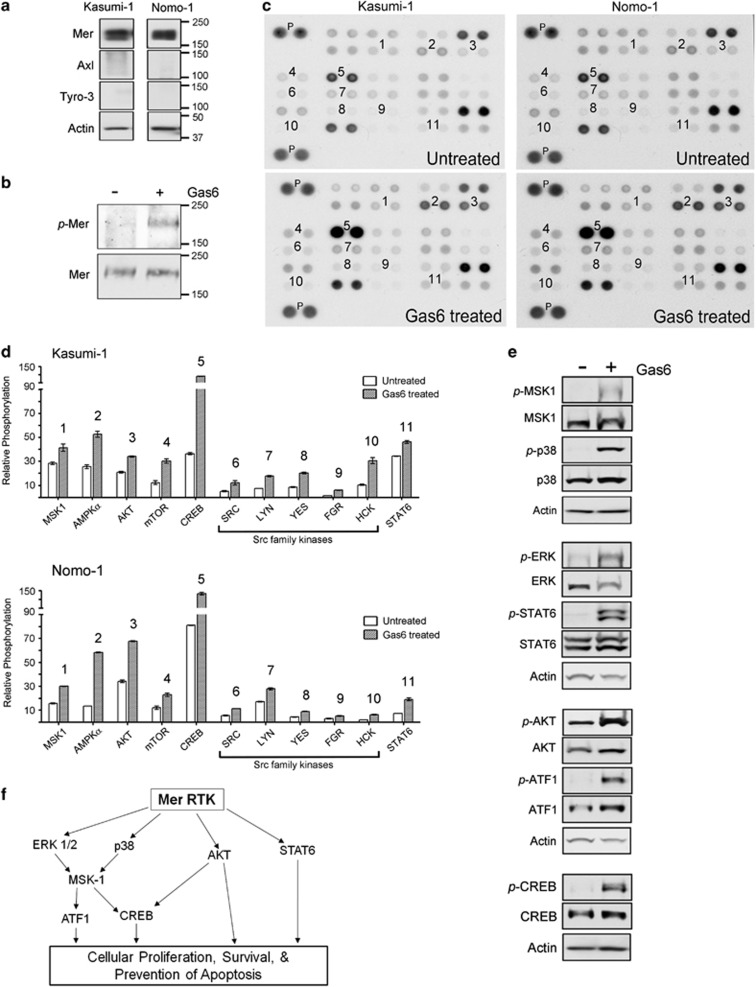
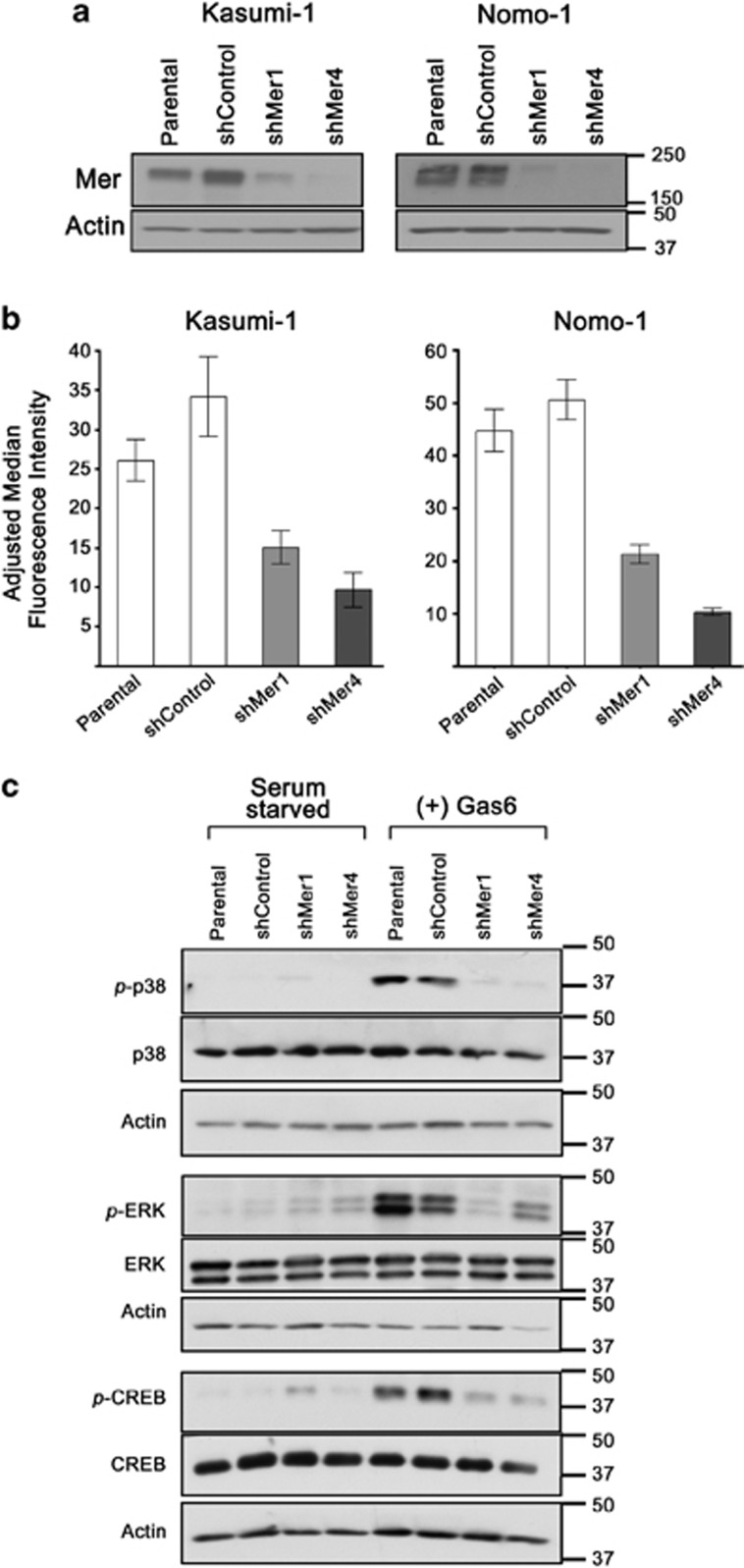
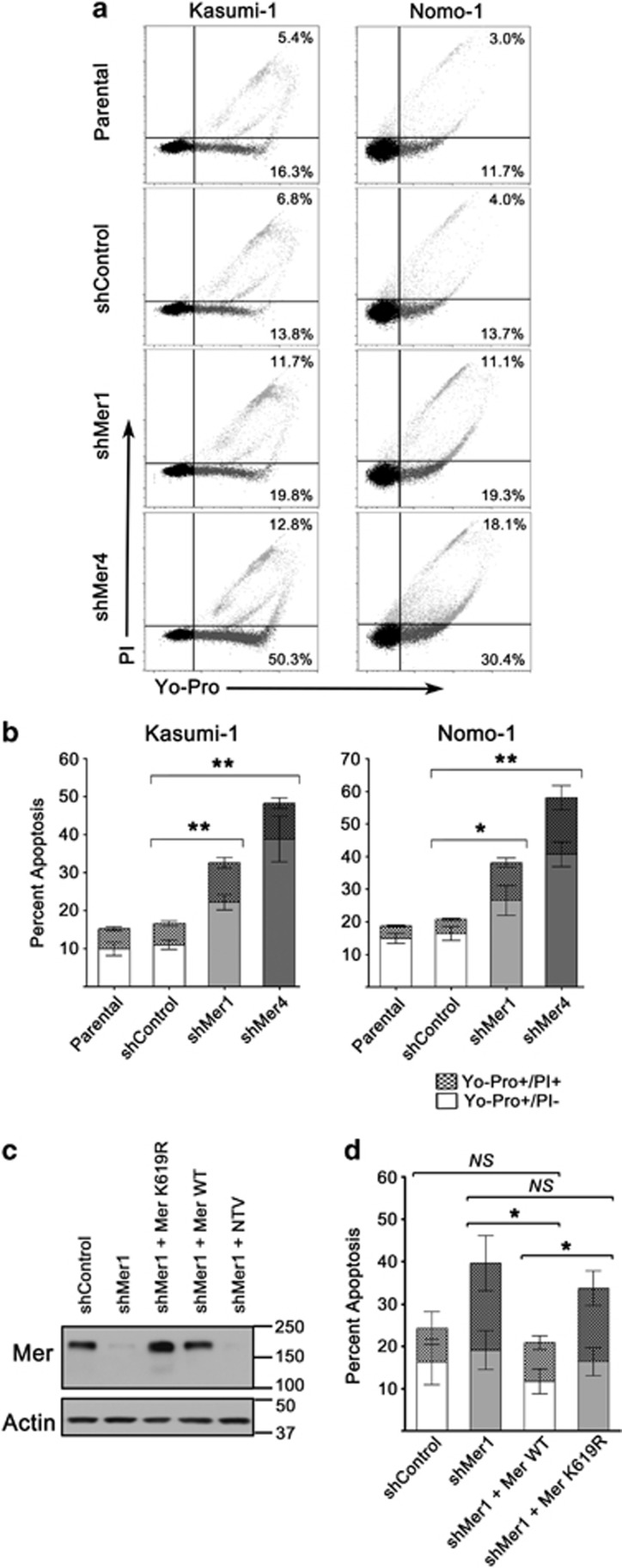
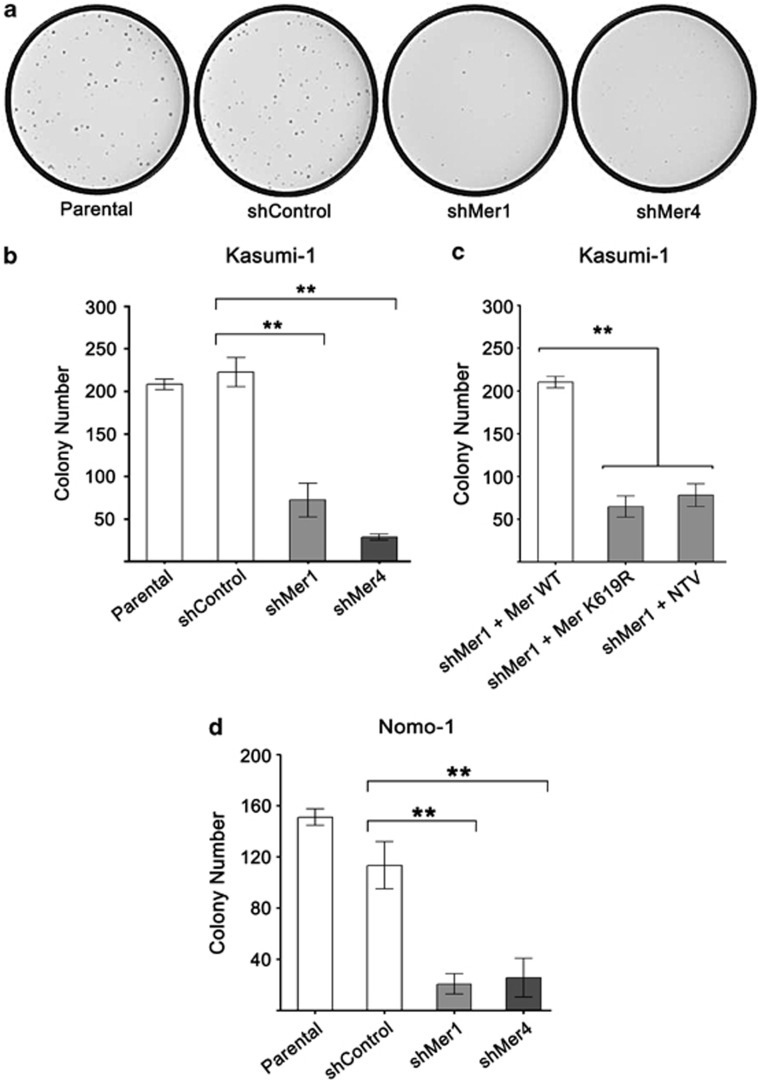
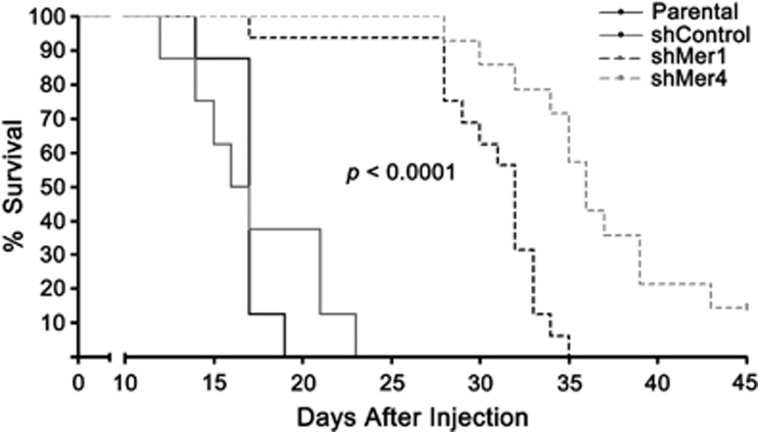
Acute myeloid leukemia (AML) continues to be extremely difficult to treat successfully, and the unacceptably low overall survival rates mandate that we assess new potential therapies to ameliorate poor clinical response to conventional therapy. Abnormal tyrosine kinase activation in AML has been associated with poor prognosis and provides strategic targets for novel therapy development. We found that Mer receptor tyrosine kinase was over-expressed in a majority of pediatric (29/36, 80%) and adult (10/10, 100%) primary AML patient blasts at the time of diagnosis, and 100% of patient samples at the time of relapse. Mer was also found to be expressed in 12 of 14 AML cell lines (86%). In contrast, normal bone marrow myeloid precursors expressed little to no Mer. Following AML cell line stimulation with Gas6, a Mer ligand, we observed activation of prosurvival and proliferative signaling pathways, including phosphorylation of ERK1/2, p38, MSK1, CREB, ATF1, AKT and STAT6. To assess the phenotypic role of Mer in AML, two independent short-hairpin RNA (shRNA) constructs were used to decrease Mer expression in the AML cell lines Nomo-1 and Kasumi-1. Reduction of Mer protein levels significantly increased rates of myeloblast apoptosis two to threefold in response to serum starvation. Furthermore, myeloblasts with knocked-down Mer demonstrated decreased colony formation by 67-87%, relative to control cell lines (P<0.01). NOD-SCID-gamma mice transplanted with Nomo-1 myeloblasts with reduced levels of Mer had a significant prolongation in survival compared with mice transplanted with the parental or control cell lines (median survival 17 days in parental and control cell lines, versus 32-36 days in Mer knockdown cell lines, P<0.0001). These data suggest a role for Mer in acute myeloid leukemogenesis and indicate that targeted inhibition of Mer may be an effective therapeutic strategy in pediatric and adult AML.
Figures
References
-
- Pulte D, Gondos A, Brenner H. Expected long-term survival of patients diagnosed with acute myeloblastic leukemia during 2006-2010. Ann Oncol. 2010;21:335–341. - PubMed
-
- Pfirrmann M, Ehninger G, Thiede C, Bornhauser M, Kramer M, Rollig C, et al. Prediction of post-remission survival in acute myeloid leukaemia: a post-hoc analysis of the AML96 trial. Lancet Oncol. 2012;13:207–214. - PubMed
-
- Sander A, Zimmermann M, Dworzak M, Fleischhack G, von Neuhoff C, Reinhardt D, et al. Consequent and intensified relapse therapy improved survival in pediatric AML: results of relapse treatment in 379 patients of three consecutive AML-BFM trials. Leukemia. 2010;24:1422–1428. - PubMed
-
- Trifilio SM, Rademaker AW, Newman D, Coyle K, Carlson-Leuer K, Mehta J, et al. Mitoxantrone and etoposide with or without intermediate dose cytarabine for the treatment of primary induction failure or relapsed acute myeloid leukemia. Leuk Res. 2012;36:394–396. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Medical
Research Materials
Miscellaneous
