

Cyclic Di-GMP modulates the disease progression of Erwinia amylovora
- PMID: 23475975
- PMCID: PMC3650540
- DOI: 10.1128/JB.02068-12
Cyclic Di-GMP modulates the disease progression of Erwinia amylovora
Erratum in
- J Bacteriol. 2013 Oct;195(20):4778
Abstract
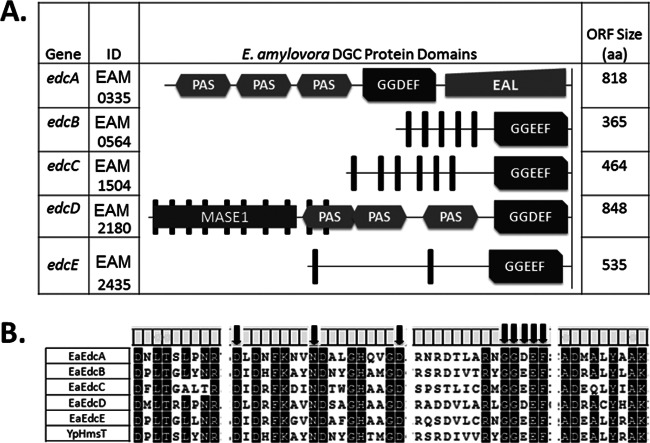
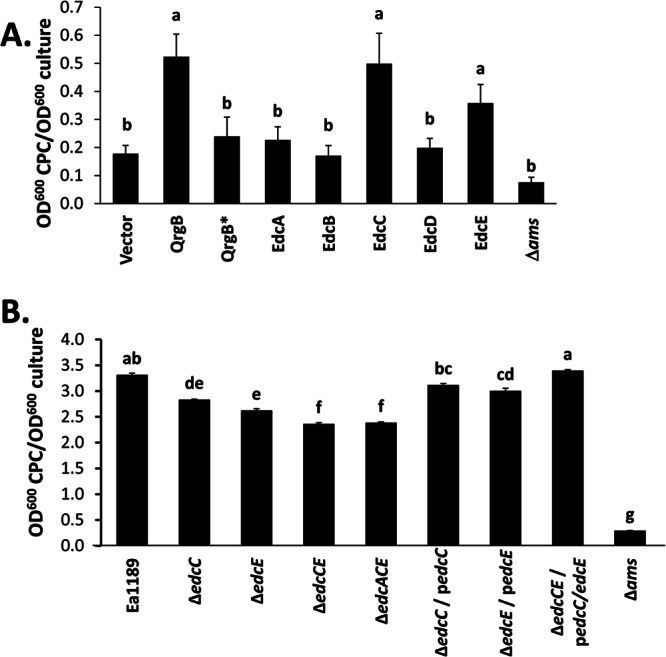
The second messenger cyclic di-GMP (c-di-GMP) is a nearly ubiquitous intracellular signal molecule known to regulate various cellular processes, including biofilm formation, motility, and virulence. The intracellular concentration of c-di-GMP is inversely governed by diguanylate cyclase (DGC) enzymes and phosphodiesterase (PDE) enzymes, which synthesize and degrade c-di-GMP, respectively. The role of c-di-GMP in the plant pathogen and causal agent of fire blight disease Erwinia amylovora has not been studied previously. Here we demonstrate that three of the five predicted DGC genes in E. amylovora (edc genes, for Erwinia diguanylate cyclase), edcA, edcC, and edcE, are active diguanylate cyclases. We show that c-di-GMP positively regulates the secretion of the main exopolysaccharide in E. amylovora, amylovoran, leading to increased biofilm formation, and negatively regulates flagellar swimming motility. Although amylovoran secretion and biofilm formation are important for the colonization of plant xylem tissues and the development of systemic infections, deletion of the two biofilm-promoting DGCs increased tissue necrosis in an immature-pear infection assay and an apple shoot infection model, suggesting that c-di-GMP negatively regulates virulence. In addition, c-di-GMP inhibited the expression of hrpA, a gene encoding the major structural component of the type III secretion pilus. Our results are the first to describe a role for c-di-GMP in E. amylovora and suggest that downregulation of motility and type III secretion by c-di-GMP during infection plays a key role in the coordination of pathogenesis.
Figures




References
-
- Eden-Green SJ, Billing E. 1974. Fireblight. Rev. Plant Pathol. 53:353–365
-
- Norelli JL, Jones AL, Aldwinckle HS. 2003. Fire blight management in the twenty-first century: using new technologies that enhance host resistance in apple. Plant Dis. 87:756–765 - PubMed
-
- Wilson M, Sigee DC, Epton HAS. 1990. Erwinia amylovora infection of hawthorn blossom. III. The nectary. J. Phytopathol. 128:62–74
-
- Wilson M, Epton HAS, Sigee DC. 1989. Erwinia amylovora infection of hawthorn blossom. II. The stigma. J. Phytopathol. 127:15–28
-
- Vanneste JL. 1995. Erwinia amylovora, p 21–46 In Singh US, Singh RP, Kohmoto K. (ed), Pathogenesis and host specificity in plant diseases: histopathological, biochemical, genetic, and molecular bases. Pergamon Press, Oxford, United Kingdom
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
