

Nkx2-5 mediates differential cardiac differentiation through interaction with Hoxa10
- PMID: 23477547
- PMCID: PMC3715789
- DOI: 10.1089/scd.2012.0611
Nkx2-5 mediates differential cardiac differentiation through interaction with Hoxa10
Abstract
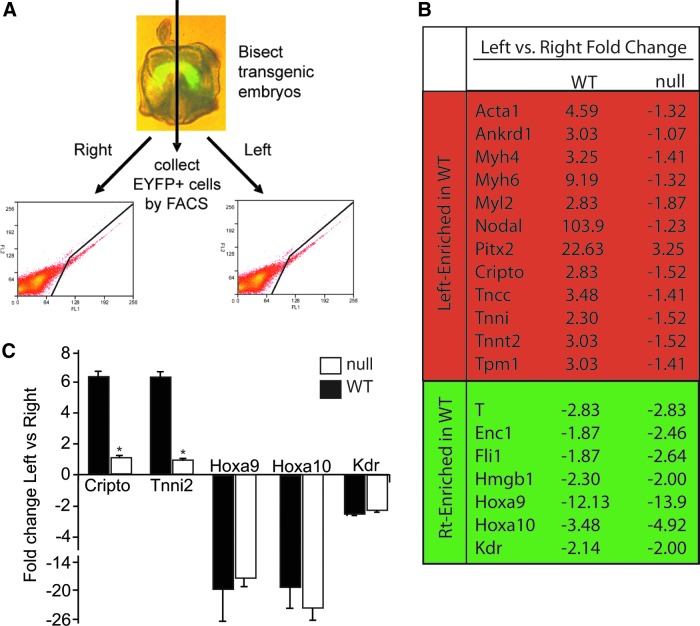
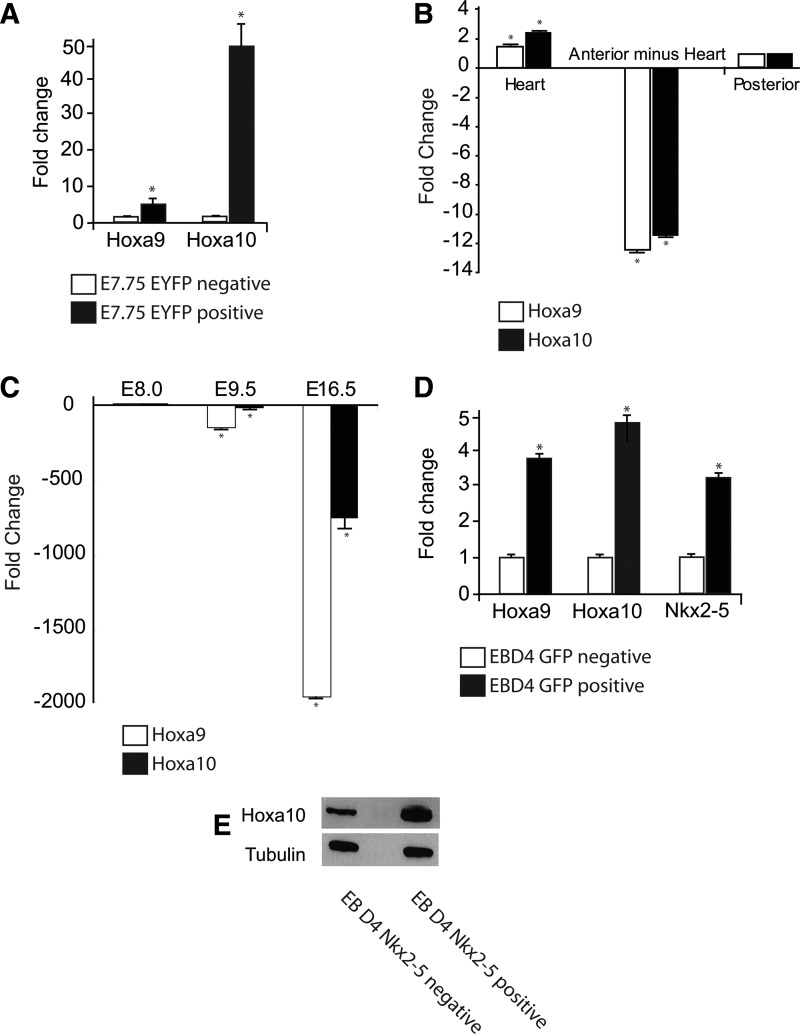
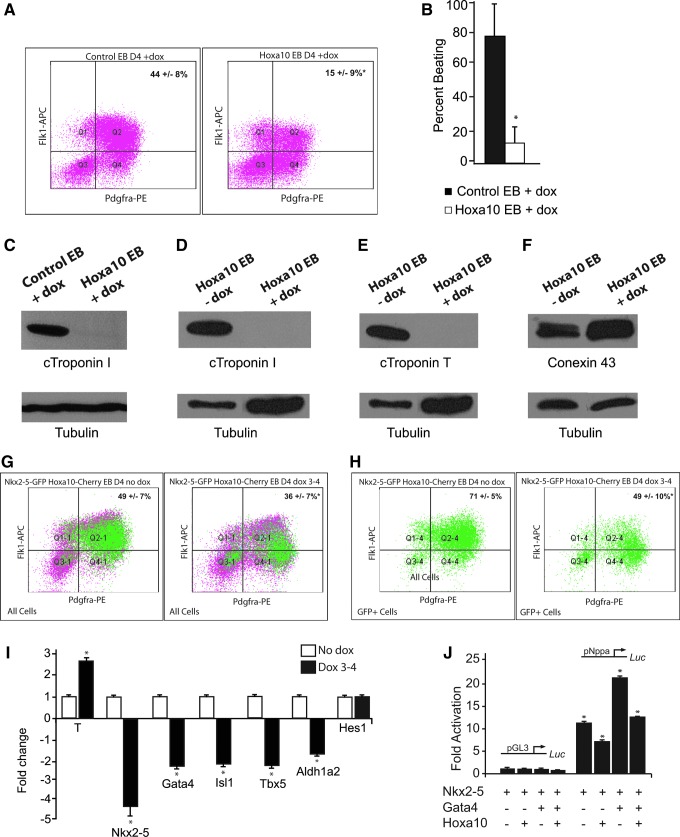
The regulation of cardiac differentiation is complex and incompletely understood. Recent studies have documented that Nkx2-5-positive cells are not limited to the cardiac lineage, but can give rise to endothelial and smooth muscle lineages. Other work has elucidated that, in addition to promoting cardiac development, Nkx2-5 plays a larger role in mesodermal patterning although the transcriptional networks that govern this developmental patterning are undefined. By profiling early Nkx2-5-positive progenitor cells, we discovered that the progenitor pools of the bisected cardiac crescent are differentiating asynchronously. This asymmetry requires Nkx2-5 as it is lost in the Nkx2-5 mutant. Surprisingly, the posterior Hox genes Hoxa9 and Hoxa10 were expressed on the right side of the cardiac crescent, independently of Nkx2-5. We describe a novel, transient, and asymmetric cardiac-specific expression pattern of the posterior Hox genes, Hoxa9 and Hoxa10, and utilize the embryonic stem cell/embryoid body (ES/EB) model system to illustrate that Hoxa10 impairs cardiac differentiation. We suggest a model whereby Hoxa10 cooperates with Nkx2-5 to regulate the timing of cardiac mesoderm differentiation.
Figures
References
-
- Hoffman JI. Incidence of congenital heart disease: II. Prenatal incidence. Pediatr Cardiol. 1995;16:155–165. - PubMed
-
- Olson EN. Srivastava D. Molecular pathways controlling heart development. Science. 1996;272:671–676. - PubMed
-
- Harvey RP. Patterning the vertebrate heart. Nat Rev Genet. 2002;3:544–556. - PubMed
-
- Olson EN. Schneider MD. Sizing up the heart: development redux in disease. Genes Dev. 2003;17:1937–1956. - PubMed
-
- Brand T. Heart development: molecular insights into cardiac specification and early morphogenesis. Dev Biol. 2003;258:1–19. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
