

Cellular effects of acute direct current stimulation: somatic and synaptic terminal effects
- PMID: 23478132
- PMCID: PMC3678043
- DOI: 10.1113/jphysiol.2012.247171
Cellular effects of acute direct current stimulation: somatic and synaptic terminal effects
Abstract
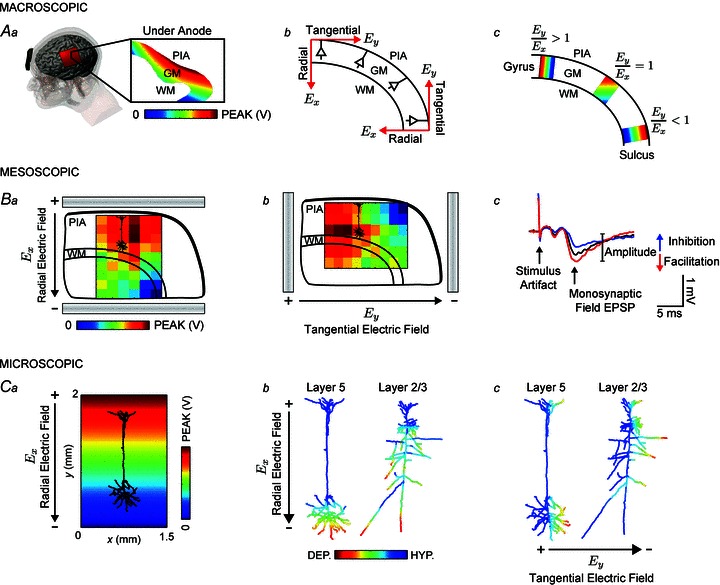
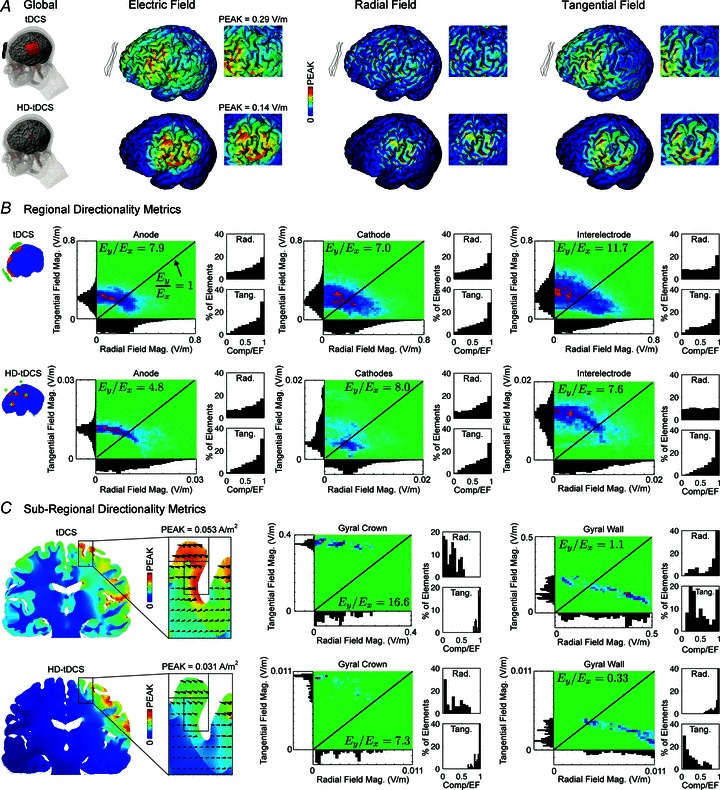
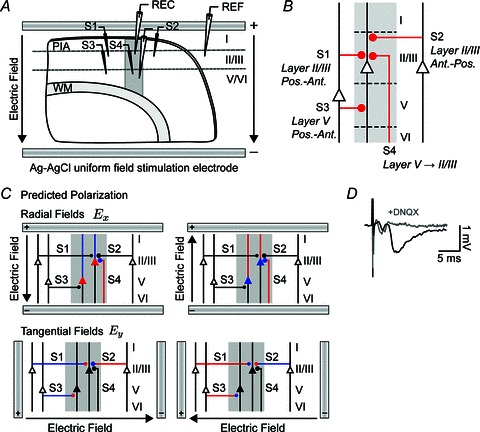
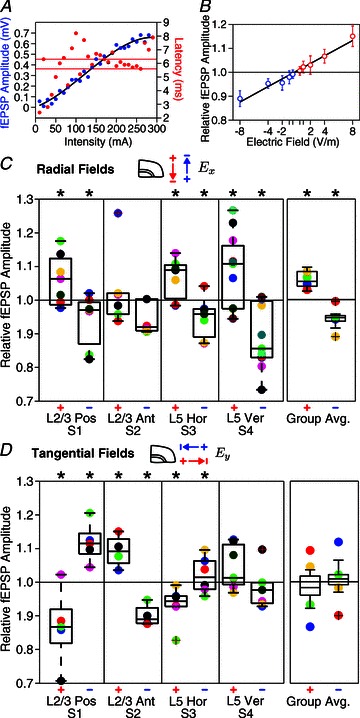
Transcranial direct current stimulation (tDCS) is a non-invasive brain stimulation technique to modulate cortical excitability. Although increased/decreased excitability under the anode/cathode electrode is nominally associated with membrane depolarization/hyperpolarization, which cellular compartments (somas, dendrites, axons and their terminals) mediate changes in cortical excitability remains unaddressed. Here we consider the acute effects of DCS on excitatory synaptic efficacy. Using multi-scale computational models and rat cortical brain slices, we show the following. (1) Typical tDCS montages produce predominantly tangential (relative to the cortical surface) direction currents (4-12 times radial direction currents), even directly under electrodes. (2) Radial current flow (parallel to the somatodendritic axis) modulates synaptic efficacy consistent with somatic polarization, with depolarization facilitating synaptic efficacy. (3) Tangential current flow (perpendicular to the somatodendritic axis) modulates synaptic efficacy acutely (during stimulation) in an afferent pathway-specific manner that is consistent with terminal polarization, with hyperpolarization facilitating synaptic efficacy. (4) Maximal polarization during uniform DCS is expected at distal (the branch length is more than three times the membrane length constant) synaptic terminals, independent of and two-three times more susceptible than pyramidal neuron somas. We conclude that during acute DCS the cellular targets responsible for modulation of synaptic efficacy are concurrently somata and axon terminals, with the direction of cortical current flow determining the relative influence.
Figures
Similar articles
-
Direct current stimulation boosts synaptic gain and cooperativity in vitro.J Physiol. 2017 Jun 1;595(11):3535-3547. doi: 10.1113/JP273005. Epub 2017 Apr 23. J Physiol. 2017. PMID: 28436038 Free PMC article.
-
Neuromodulation of Axon Terminals.Cereb Cortex. 2018 Aug 1;28(8):2786-2794. doi: 10.1093/cercor/bhx158. Cereb Cortex. 2018. PMID: 28655149 Free PMC article.
-
Multi-scale model of axonal and dendritic polarization by transcranial direct current stimulation in realistic head geometry.Brain Stimul. 2023 Nov-Dec;16(6):1776-1791. doi: 10.1016/j.brs.2023.11.018. Epub 2023 Dec 6. Brain Stimul. 2023. PMID: 38056825 Free PMC article.
-
Animal models of transcranial direct current stimulation: Methods and mechanisms.Clin Neurophysiol. 2016 Nov;127(11):3425-3454. doi: 10.1016/j.clinph.2016.08.016. Epub 2016 Sep 10. Clin Neurophysiol. 2016. PMID: 27693941 Free PMC article. Review.
-
Backpropagation of action potentials generated at ectopic axonal loci: hypothesis that axon terminals integrate local environmental signals.Brain Res Brain Res Rev. 1995 Jul;21(1):42-92. doi: 10.1016/0165-0173(95)00004-m. Brain Res Brain Res Rev. 1995. PMID: 8547954 Review.
Cited by
-
Slow-Frequency Pulsed Transcranial Electrical Stimulation for Modulation of Cortical Plasticity Based on Reciprocity Targeting with Precision Electrical Head Modeling.Front Hum Neurosci. 2016 Aug 2;10:377. doi: 10.3389/fnhum.2016.00377. eCollection 2016. Front Hum Neurosci. 2016. PMID: 27531976 Free PMC article.
-
Transcranial Direct Current Stimulation Enhances Exercise Performance: A Mini Review of the Underlying Mechanisms.Front Neuroergon. 2022 Apr 26;3:841911. doi: 10.3389/fnrgo.2022.841911. eCollection 2022. Front Neuroergon. 2022. PMID: 38235480 Free PMC article. Review.
-
Perspectives on the Combined Use of Electric Brain Stimulation and Perceptual Learning in Vision.Vision (Basel). 2022 Jun 14;6(2):33. doi: 10.3390/vision6020033. Vision (Basel). 2022. PMID: 35737420 Free PMC article.
-
Safety and efficacy of adjunctive transcranial direct current stimulation in treatment-resistant obsessive-compulsive disorder: An open-label trial.Indian J Psychiatry. 2019;61(4):327-334. doi: 10.4103/psychiatry.IndianJPsychiatry_509_18. Indian J Psychiatry. 2019. PMID: 31391634 Free PMC article.
-
Current challenges: the ups and downs of tACS.Exp Brain Res. 2019 Dec;237(12):3071-3088. doi: 10.1007/s00221-019-05666-0. Epub 2019 Oct 16. Exp Brain Res. 2019. PMID: 31620829 Review.
References
-
- Andreasen M, Nedergaard S. Dendritic electrogenesis in rat hippocampal CA1 pyramidal neurons: functional aspects of Na+ and Ca2+ currents in apical dendrites. Hippocampus. 1996;6:79–95. - PubMed
-
- Antal A, Varga ET, Kincses TZ, Nitsche MA, Paulus W. Oscillatory brain activity and transcranial direct current stimulation in humans. Neuroreport. 2004;15:1307–1310. - PubMed
-
- Arlotti M, Rahman A, Minhas P, Bikson M. Axon terminal polarization induced by weak uniform DC electric fields: a modeling study. Conf Proc IEEE Eng Med Biol Soc. 2012;2012:4575–4578. - PubMed
-
- Aroniadou VA, Keller A. The patterns and synaptic properties of horizontal intracortical connections in the rat motor cortex. J Neurophysiol. 1993;70:1553–1569. - PubMed
Publication types
MeSH terms
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources