

Distinct phospholipase C-β isozymes mediate lysophosphatidic acid receptor 1 effects on intestinal epithelial homeostasis and wound closure
- PMID: 23478264
- PMCID: PMC3647975
- DOI: 10.1128/MCB.00038-13
Distinct phospholipase C-β isozymes mediate lysophosphatidic acid receptor 1 effects on intestinal epithelial homeostasis and wound closure
Abstract
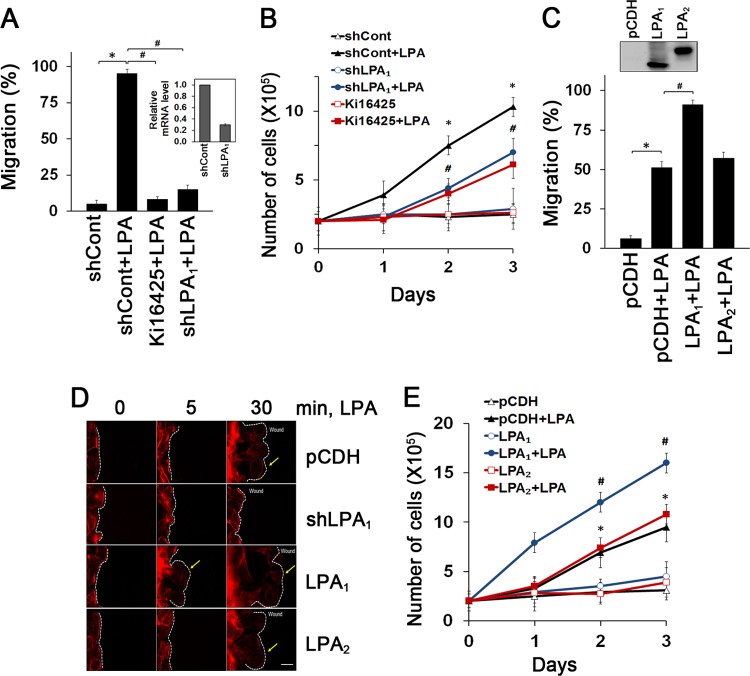
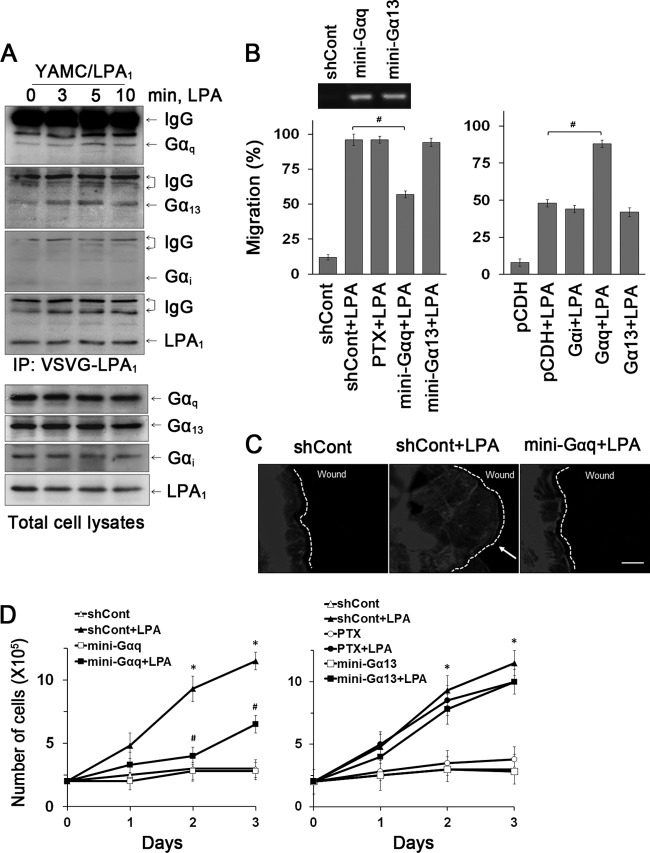
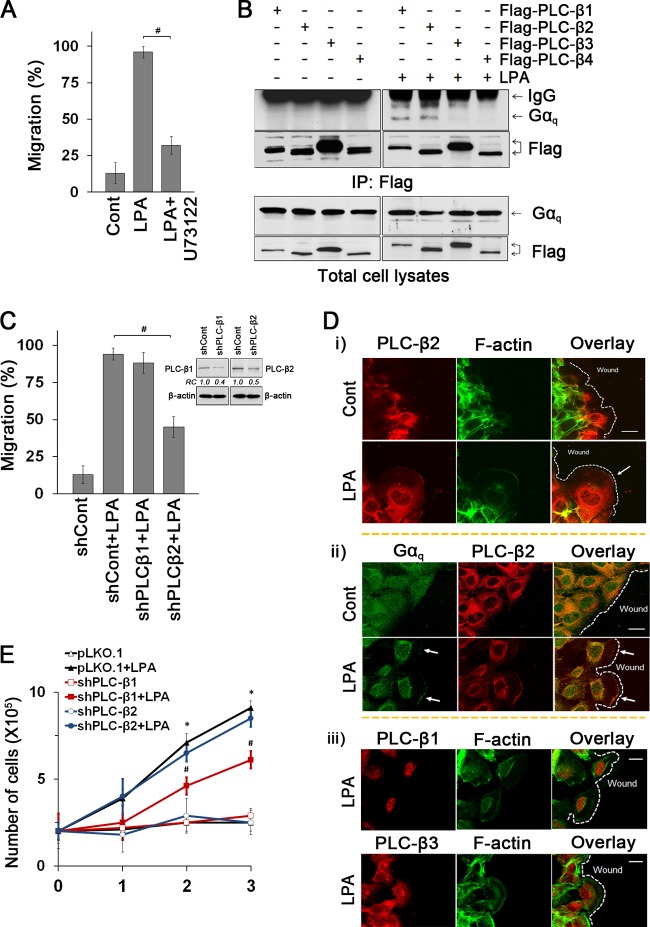
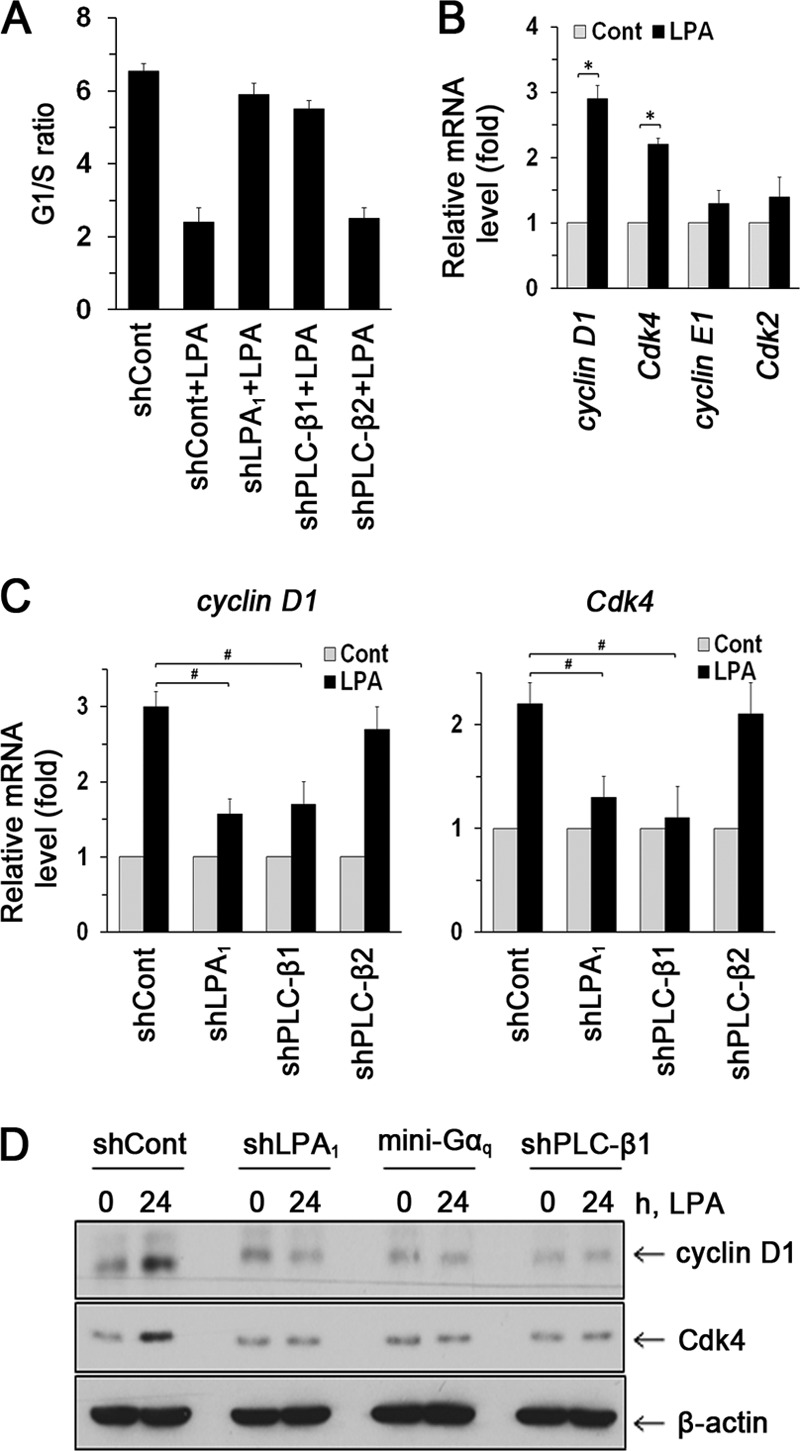
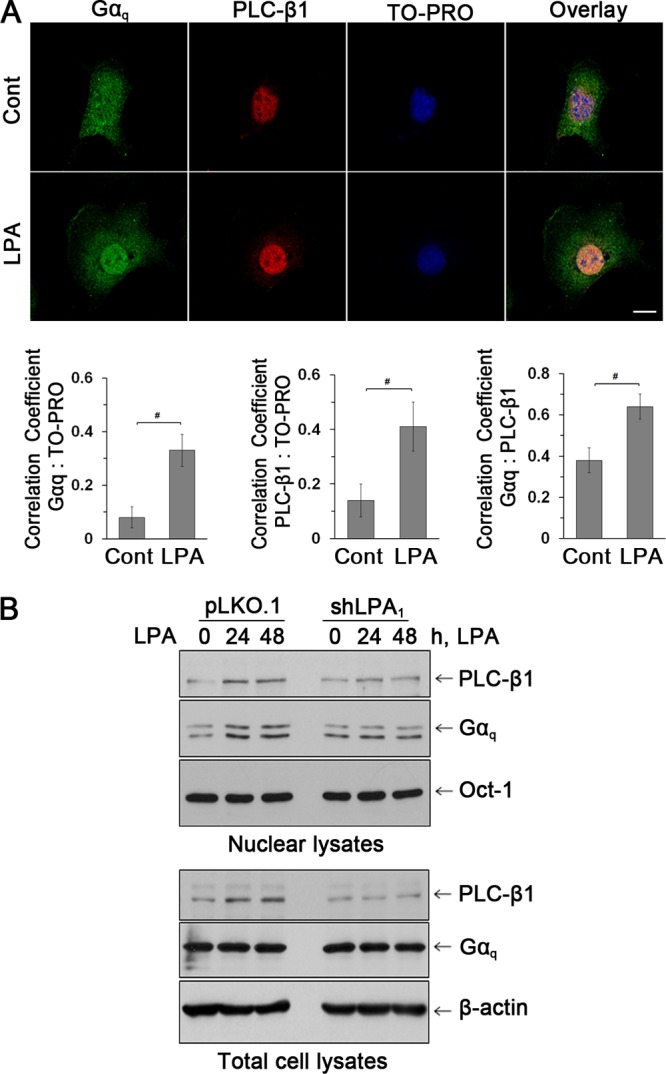
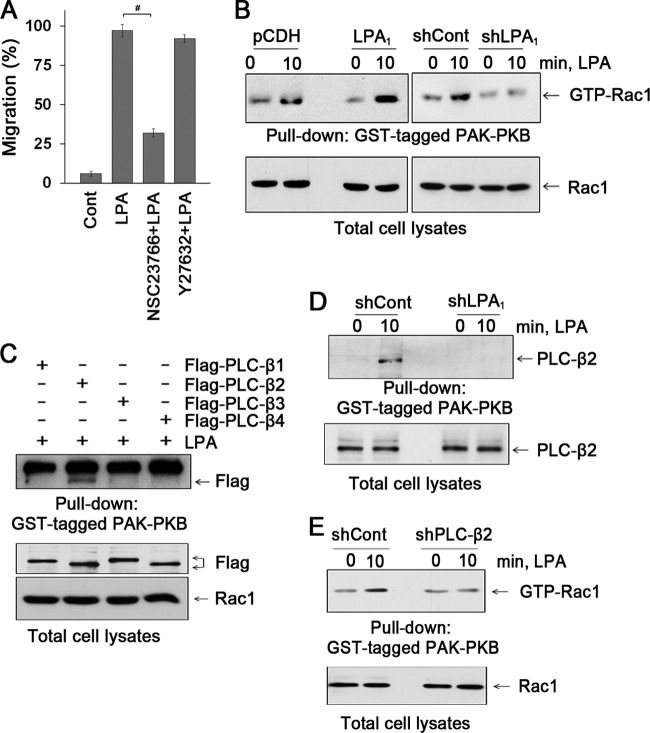
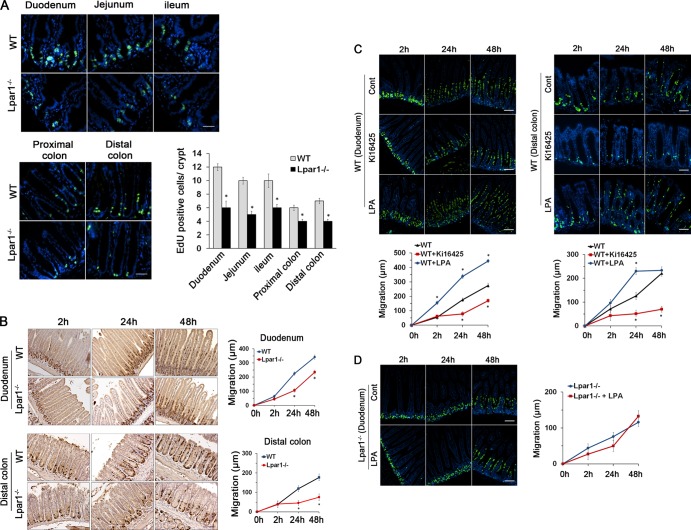
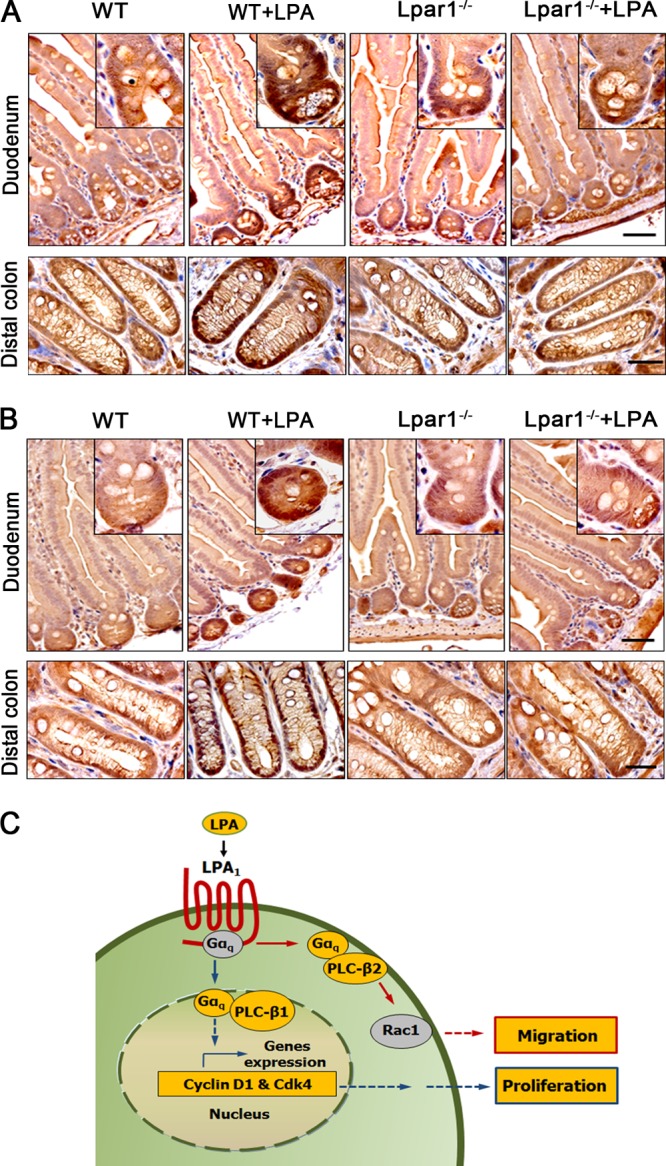
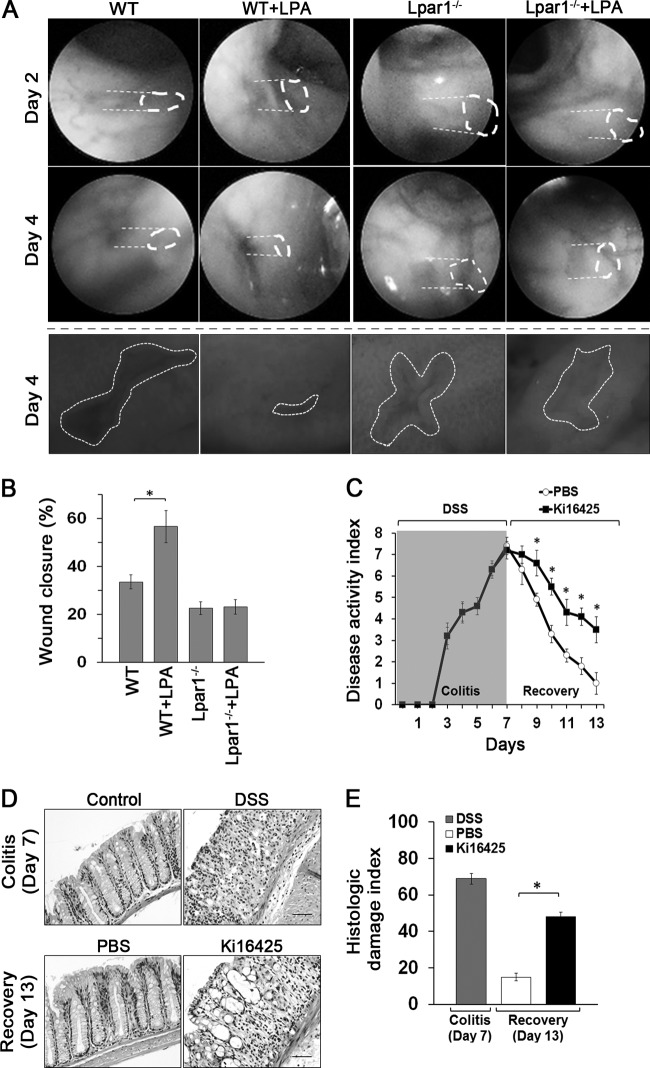
Maintenance of the epithelial barrier in the intestinal tract is necessary to protect the host from the hostile luminal environment. Phospholipase C-β (PLC-β) has been implicated to control myriad signaling cascades. However, the biological effects of selective PLC-β isozymes are poorly understood. We describe novel findings that lysophosphatidic acid (LPA) regulates PLC-β1 and PLC-β2 via two distinct pathways to enhance intestinal epithelial cell (IEC) proliferation and migration that facilitate wound closure and recovery of the intestinal epithelial barrier. LPA acting on the LPA1 receptor promotes IEC migration by facilitating the interaction of Gαq with PLC-β2. LPA-induced cell proliferation is PLC-β1 dependent and involves translocation of Gαq to the nucleus, where it interacts with PLC-β1 to induce cell cycle progression. An in vivo study using LPA1-deficient mice (Lpar1(-/-)) shows a decreased number of proliferating IECs and migration along the crypt-luminal axis. Additionally, LPA enhances migration and proliferation of IECs in an LPA1-dependent manner, and Lpar1(-/-) mice display defective mucosal wound repair that requires cell proliferation and migration. These findings delineate novel LPA1-dependent lipid signaling that facilitates mucosal wound repair via spatial targeting of distinct PLC-βs within the cell.
Figures
References
-
- Crosnier C, Stamataki D, Lewis J. 2006. Organizing cell renewal in the intestine: stem cells, signals and combinatorial control. Nat. Rev. Genet. 7:349–359 - PubMed
-
- Choi JW, Herr DR, Noguchi K, Yung YC, Lee CW, Mutoh T, Lin ME, Teo ST, Park KE, Mosley AN, Chun J. 2010. LPA receptors: subtypes and biological actions. Annu. Rev. Pharmacol. Toxicol. 50:157–186 - PubMed
-
- Sturm A, Sudermann T, Schulte KM, Goebell H, Dignass AU. 1999. Modulation of intestinal epithelial wound healing in vitro and in vivo by lysophosphatidic acid. Gastroenterology 117:368–377 - PubMed
-
- Lin S, Yeruva S, He P, Singh AK, Zhang H, Chen M, Lamprecht G, de Jonge HR, Tse M, Donowitz M, Hogema BM, Chun J, Seidler U, Yun CC. 2010. Lysophosphatidic acid stimulates the intestinal brush border Na(+)/H(+) exchanger 3 and fluid absorption via LPA(5) and NHERF2. Gastroenterology 138:649–658 - PMC - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
Miscellaneous