

Splicing-factor oncoprotein SRSF1 stabilizes p53 via RPL5 and induces cellular senescence
- PMID: 23478443
- PMCID: PMC3628402
- DOI: 10.1016/j.molcel.2013.02.001
Splicing-factor oncoprotein SRSF1 stabilizes p53 via RPL5 and induces cellular senescence
Abstract
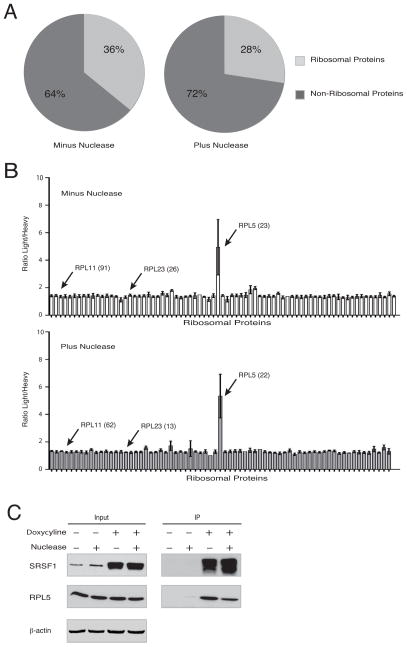
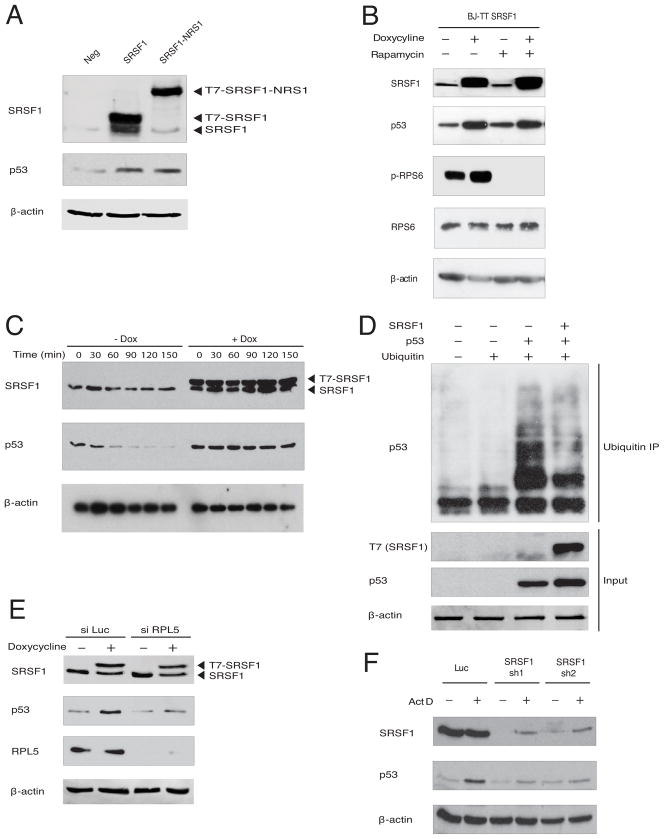
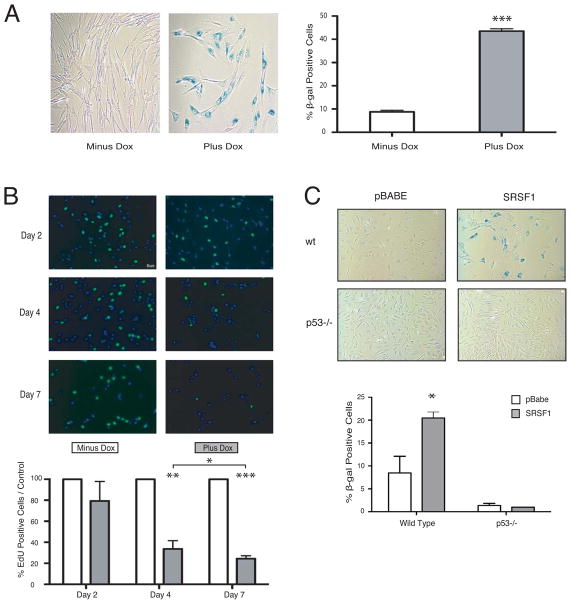
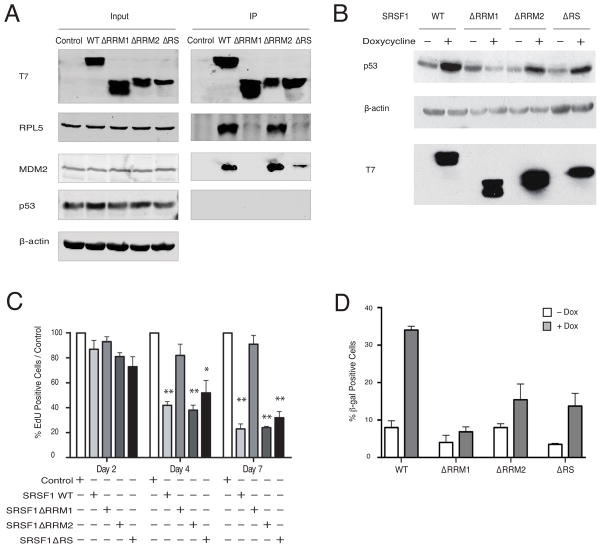
Splicing and translation are highly regulated steps of gene expression. Altered expression of proteins involved in these processes can be deleterious. Therefore, the cell has many safeguards against such misregulation. We report that the oncogenic splicing factor SRSF1, which is overexpressed in many cancers, stabilizes the tumor suppressor protein p53 by abrogating its MDM2-dependent proteasomal degradation. We show that SRSF1 is a necessary component of an MDM2/ribosomal protein complex, separate from the ribosome, that functions in a p53-dependent ribosomal-stress checkpoint pathway. Consistent with the stabilization of p53, increased SRSF1 expression in primary human fibroblasts decreases cellular proliferation and ultimately triggers oncogene-induced senescence (OIS). These findings underscore the deleterious outcome of SRSF1 overexpression and identify a cellular defense mechanism against its aberrant function. Furthermore, they implicate the RPL5-MDM2 complex in OIS and demonstrate a link between spliceosomal and ribosomal components, functioning independently of their canonical roles, to monitor cellular physiology and cell-cycle progression.
Copyright © 2013 Elsevier Inc. All rights reserved.
Figures


Comment in
-
A new path to oncogene-induced senescence: at the crossroads of splicing and translation.Cell Cycle. 2013 May 15;12(10):1477-9. doi: 10.4161/cc.24749. Epub 2013 Apr 22. Cell Cycle. 2013. PMID: 23624837 Free PMC article. No abstract available.
Similar articles
-
A new path to oncogene-induced senescence: at the crossroads of splicing and translation.Cell Cycle. 2013 May 15;12(10):1477-9. doi: 10.4161/cc.24749. Epub 2013 Apr 22. Cell Cycle. 2013. PMID: 23624837 Free PMC article. No abstract available.
-
Perturbation of RNA Polymerase I transcription machinery by ablation of HEATR1 triggers the RPL5/RPL11-MDM2-p53 ribosome biogenesis stress checkpoint pathway in human cells.Cell Cycle. 2018;17(1):92-101. doi: 10.1080/15384101.2017.1403685. Epub 2017 Dec 10. Cell Cycle. 2018. PMID: 29143558 Free PMC article.
-
Ribosomal protein L4 is a novel regulator of the MDM2-p53 loop.Oncotarget. 2016 Mar 29;7(13):16217-26. doi: 10.18632/oncotarget.7479. Oncotarget. 2016. PMID: 26908445 Free PMC article.
-
Mdm2 widens its repertoire.Cell Cycle. 2007 Apr 1;6(7):827-9. doi: 10.4161/cc.6.7.4086. Epub 2007 Apr 28. Cell Cycle. 2007. PMID: 17377491 Review.
-
The Evolution of the Ribosomal Protein-MDM2-p53 Pathway.Cold Spring Harb Perspect Med. 2016 Dec 1;6(12):a026138. doi: 10.1101/cshperspect.a026138. Cold Spring Harb Perspect Med. 2016. PMID: 27908926 Free PMC article. Review.
Cited by
-
The Central Role of Ribosomal Proteins in p53 Regulation.Cancers (Basel). 2025 May 8;17(10):1597. doi: 10.3390/cancers17101597. Cancers (Basel). 2025. PMID: 40427096 Free PMC article.
-
LncRNA BBOX1-AS1 Contributes to Laryngeal Carcinoma Progression by Recruiting SRSF1 to Maintain EFNB2 mRNA Stability.Biochem Genet. 2025 Aug;63(4):3380-3398. doi: 10.1007/s10528-024-10879-2. Epub 2024 Jul 4. Biochem Genet. 2025. PMID: 38965134
-
Differential connectivity of splicing activators and repressors to the human spliceosome.Genome Biol. 2015 Jun 6;16(1):119. doi: 10.1186/s13059-015-0682-5. Genome Biol. 2015. PMID: 26047612 Free PMC article.
-
RNA binding proteins (RBPs) and their role in DNA damage and radiation response in cancer.Adv Drug Deliv Rev. 2022 Dec;191:114569. doi: 10.1016/j.addr.2022.114569. Epub 2022 Oct 14. Adv Drug Deliv Rev. 2022. PMID: 36252617 Free PMC article. Review.
-
Partial least squares based gene expression analysis in renal failure.Diagn Pathol. 2014 Jul 5;9:137. doi: 10.1186/1746-1596-9-137. Diagn Pathol. 2014. PMID: 24997640 Free PMC article.
References
-
- Bartkova J, Rezaei N, Liontos M, Karakaidos P, Kletsas D, Issaeva N, Vassiliou LV, Kolettas E, Niforou K, Zoumpourlis VC, et al. Oncogene-induced senescence is part of the tumorigenesis barrier imposed by DNA damage checkpoints. Nature. 2006;444:633–637. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
Research Materials
Miscellaneous
