

Cdc48-associated complex bound to 60S particles is required for the clearance of aberrant translation products
- PMID: 23479637
- PMCID: PMC3612664
- DOI: 10.1073/pnas.1221724110
Cdc48-associated complex bound to 60S particles is required for the clearance of aberrant translation products
Abstract
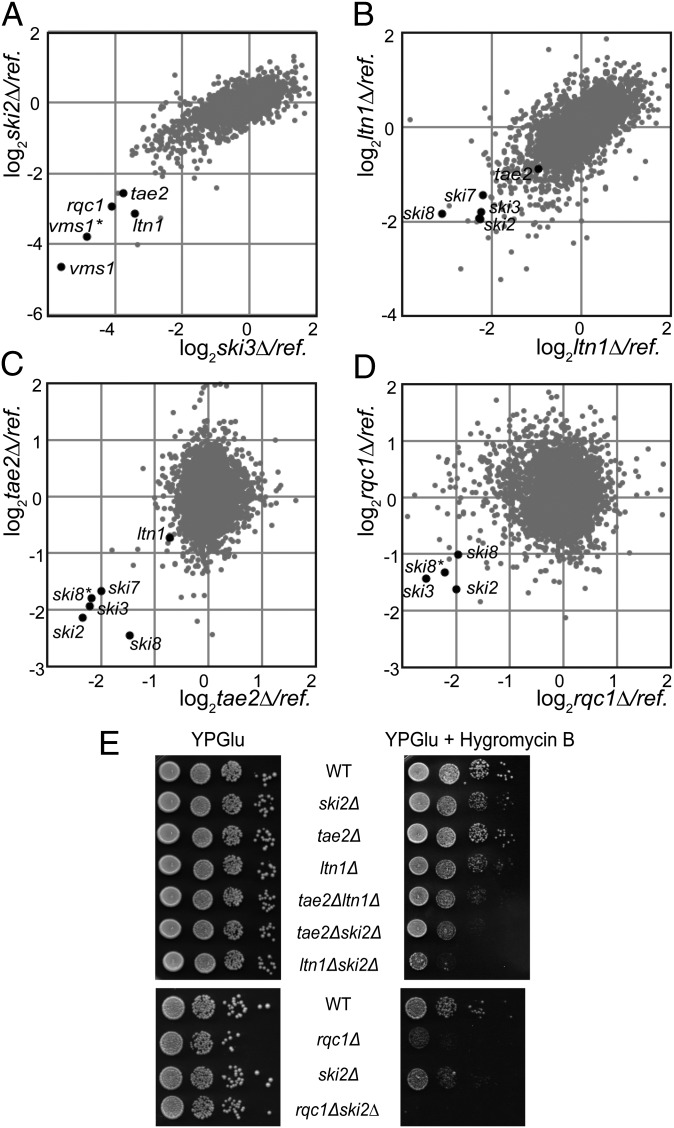
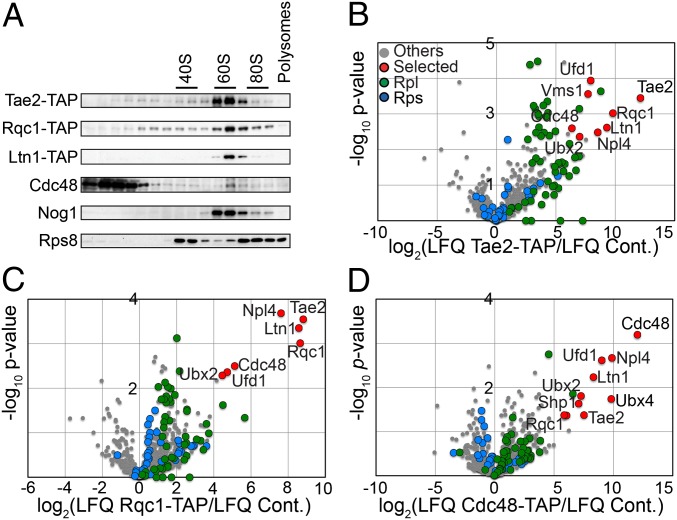
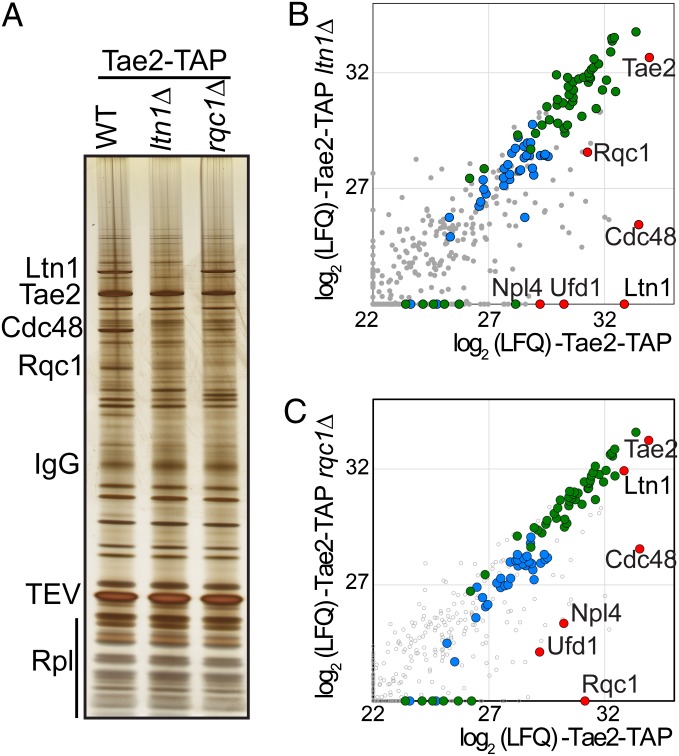
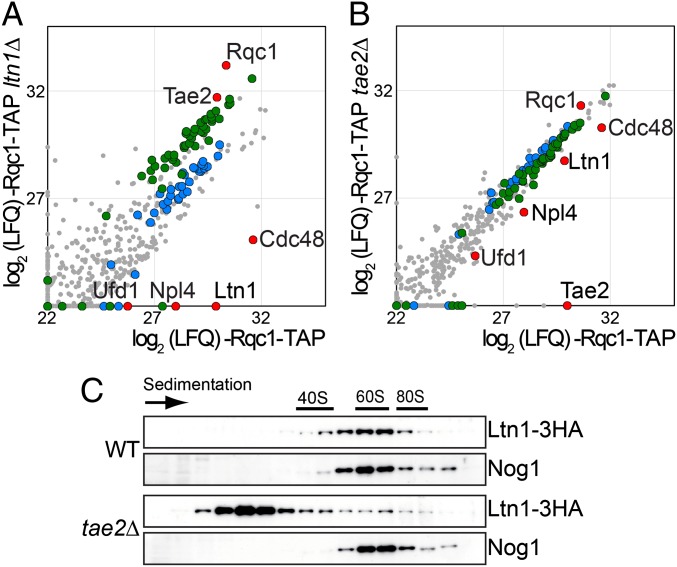
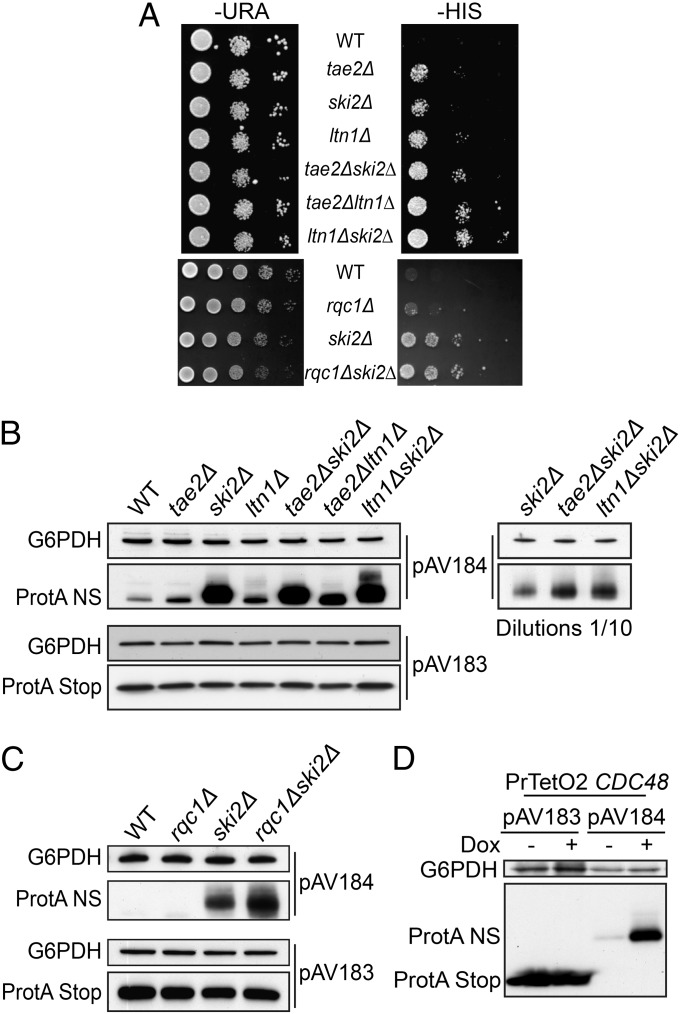
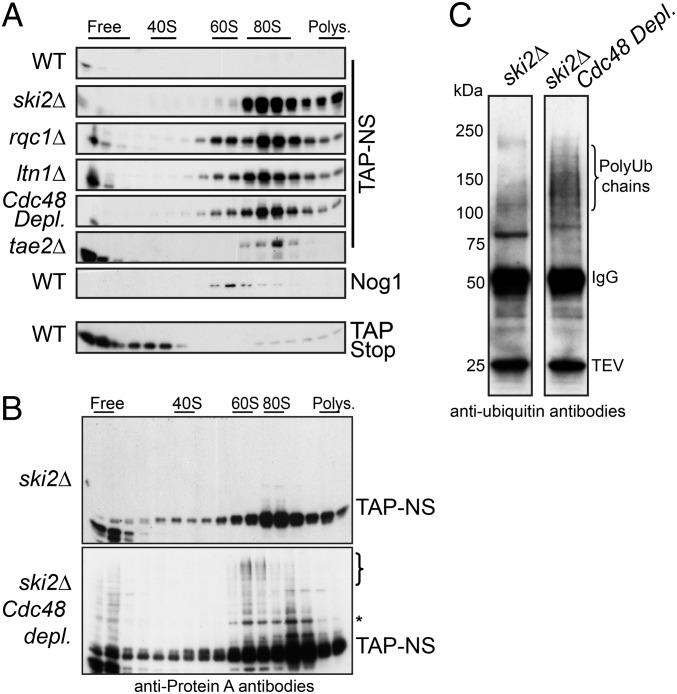
Ribosome stalling on eukaryotic mRNAs triggers cotranslational RNA and protein degradation through conserved mechanisms. For example, mRNAs lacking a stop codon are degraded by the exosome in association with its cofactor, the SKI complex, whereas the corresponding aberrant nascent polypeptides are ubiquitinated by the E3 ligases Ltn1 and Not4 and become proteasome substrates. How translation arrest is linked with polypeptide degradation is still unclear. Genetic screens with SKI and LTN1 mutants allowed us to identify translation-associated element 2 (Tae2) and ribosome quality control 1 (Rqc1), two factors that we found associated, together with Ltn1 and the AAA-ATPase Cdc48, to 60S ribosomal subunits. Translation-associated element 2 (Tae2), Rqc1, and Cdc48 were all required for degradation of polypeptides synthesized from Non-Stop mRNAs (Non-Stop protein decay; NSPD). Both Ltn1 and Rqc1 were essential for the recruitment of Cdc48 to 60S particles. Polysome gradient analyses of mutant strains revealed unique intermediates of this pathway, showing that the polyubiquitination of Non-Stop peptides is a progressive process. We propose that ubiquitination of the nascent peptide starts on the 80S and continues on the 60S, on which Cdc48 is recruited to escort the substrate for proteasomal degradation.
Conflict of interest statement
The authors declare no conflict of interest.
Figures
References
-
- Frischmeyer PA, et al. (2002) An mRNA surveillance mechanism that eliminates transcripts lacking termination codons. Science 295:2258–2261. - PubMed
-
- van Hoof A, Frischmeyer PA, Dietz HC, Parker R. Exosome-mediated recognition and degradation of mRNAs lacking a termination codon. Science. 2002;295(5563):2262–2264. - PubMed
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
