

An electron-bifurcating caffeyl-CoA reductase
- PMID: 23479729
- PMCID: PMC3630892
- DOI: 10.1074/jbc.M112.444919
An electron-bifurcating caffeyl-CoA reductase
Abstract
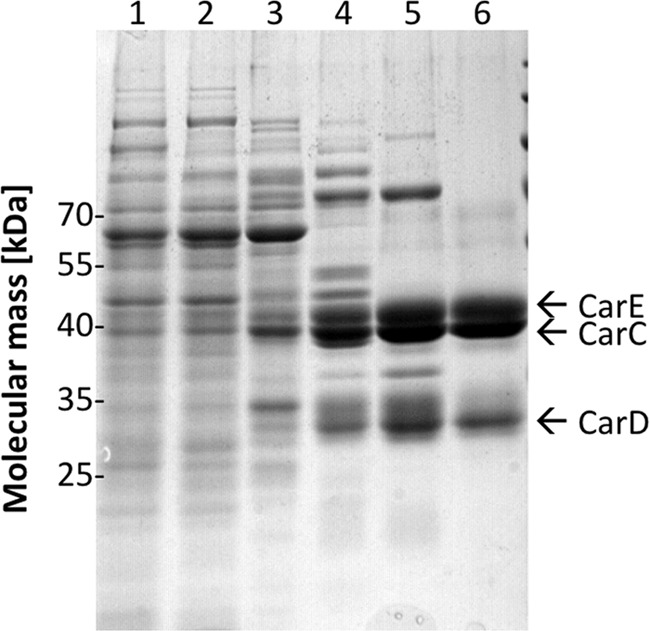
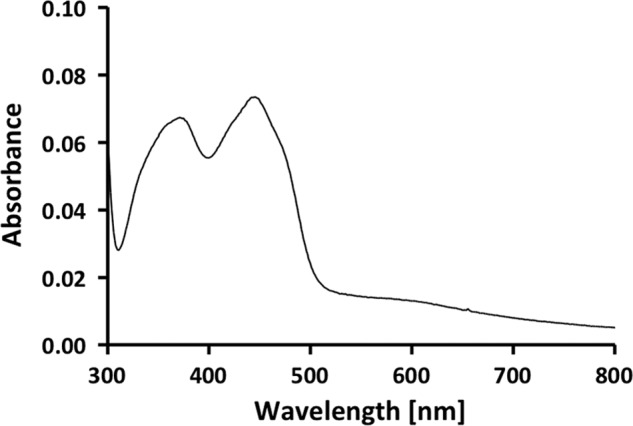
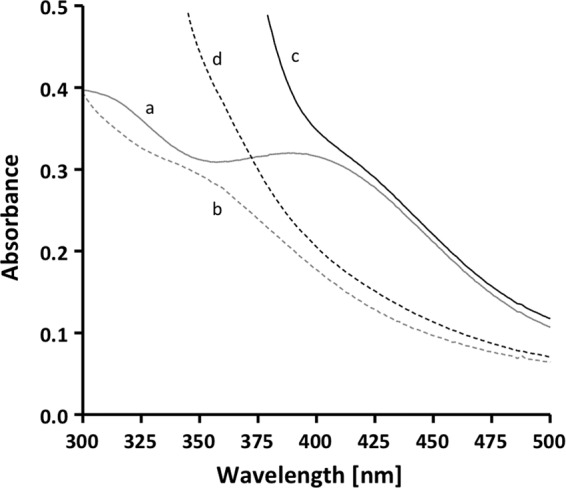
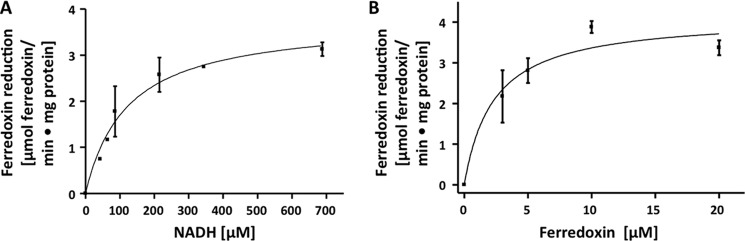
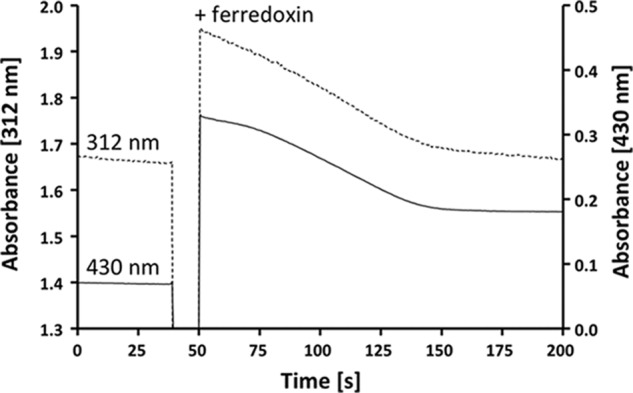
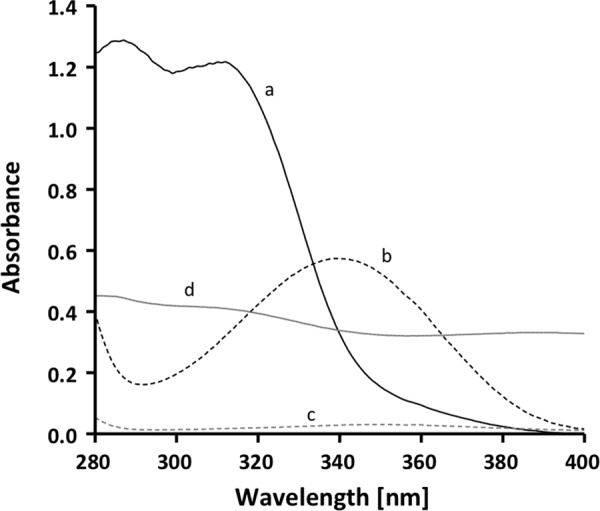
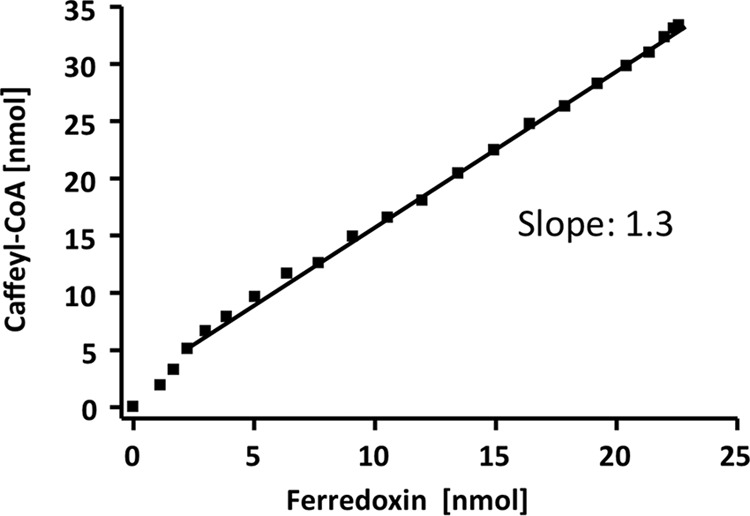
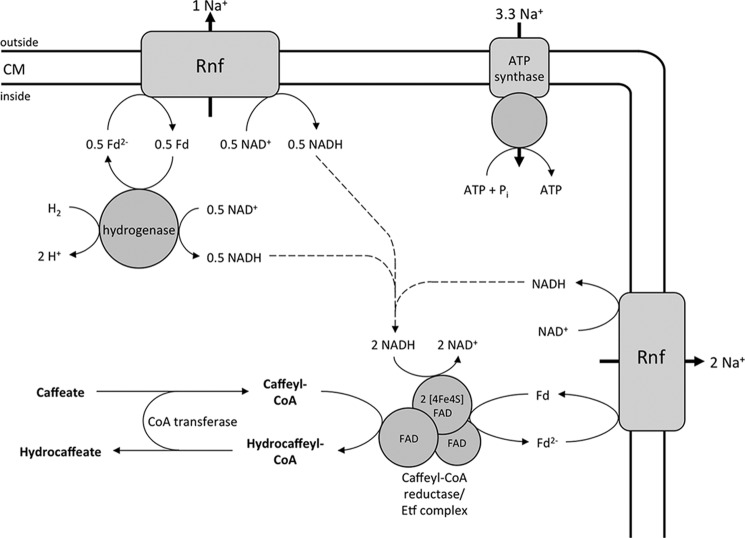
A low potential electron carrier ferredoxin (E0' ≈ -500 mV) is used to fuel the only bioenergetic coupling site, a sodium-motive ferredoxin:NAD(+) oxidoreductase (Rnf) in the acetogenic bacterium Acetobacterium woodii. Because ferredoxin reduction with physiological electron donors is highly endergonic, it must be coupled to an exergonic reaction. One candidate is NADH-dependent caffeyl-CoA reduction. We have purified a complex from A. woodii that contains a caffeyl-CoA reductase and an electron transfer flavoprotein. The enzyme contains three subunits encoded by the carCDE genes and is predicted to have, in addition to FAD, two [4Fe-4S] clusters as cofactor, which is consistent with the experimental determination of 4 mol of FAD, 9 mol of iron, and 9 mol of acid-labile sulfur. The enzyme complex catalyzed caffeyl-CoA-dependent oxidation of reduced methyl viologen. With NADH as donor, it catalyzed caffeyl-CoA reduction, but this reaction was highly stimulated by the addition of ferredoxin. Spectroscopic analyses revealed that ferredoxin and caffeyl-CoA were reduced simultaneously, and a stoichiometry of 1.3:1 was determined. Apparently, the caffeyl-CoA reductase-Etf complex of A. woodii uses the novel mechanism of flavin-dependent electron bifurcation to drive the endergonic ferredoxin reduction with NADH as reductant by coupling it to the exergonic NADH-dependent reduction of caffeyl-CoA.
Figures
References
-
- Müller V. (2008) Bacterial fermentation. in Encyclopedia of Life Sciences, John Wiley & Sons Ltd., Chichester, United Kingdom, DOI: 10.1002/9780470015902.a0001415.pub2 - DOI
-
- Schmitz R. A., Daniel R., Deppenmeier U., Gottschalk G. (2006) The anaerobic way of life. in The Prokaryotes (Dworkin M., Falkow S., Rosenberg E., Schleifer K. H., Stackebrandt E., eds) Vol. 2, pp. 86–101, Springer-Verlag, New York
-
- Dimroth P., von Ballmoos C. (2008) ATP synthesis by decarboxylation phosphorylation. Results Probl. Cell Differ. 45, 153–184 - PubMed
-
- Gottschalk G., Thauer R. K. (2001) The Na+-translocating methyltransferase complex from methanogenic archaea. Biochim. Biophys. Acta 1505, 28–36 - PubMed
-
- Buckel W. (2001) Sodium ion-translocating decarboxylases. Biochim. Biophys. Acta 1505, 15–27 - PubMed
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
