

Deletion of autophagy-related 5 (Atg5) and Pik3c3 genes in the lens causes cataract independent of programmed organelle degradation
- PMID: 23479732
- PMCID: PMC3630873
- DOI: 10.1074/jbc.M112.437103
Deletion of autophagy-related 5 (Atg5) and Pik3c3 genes in the lens causes cataract independent of programmed organelle degradation
Abstract
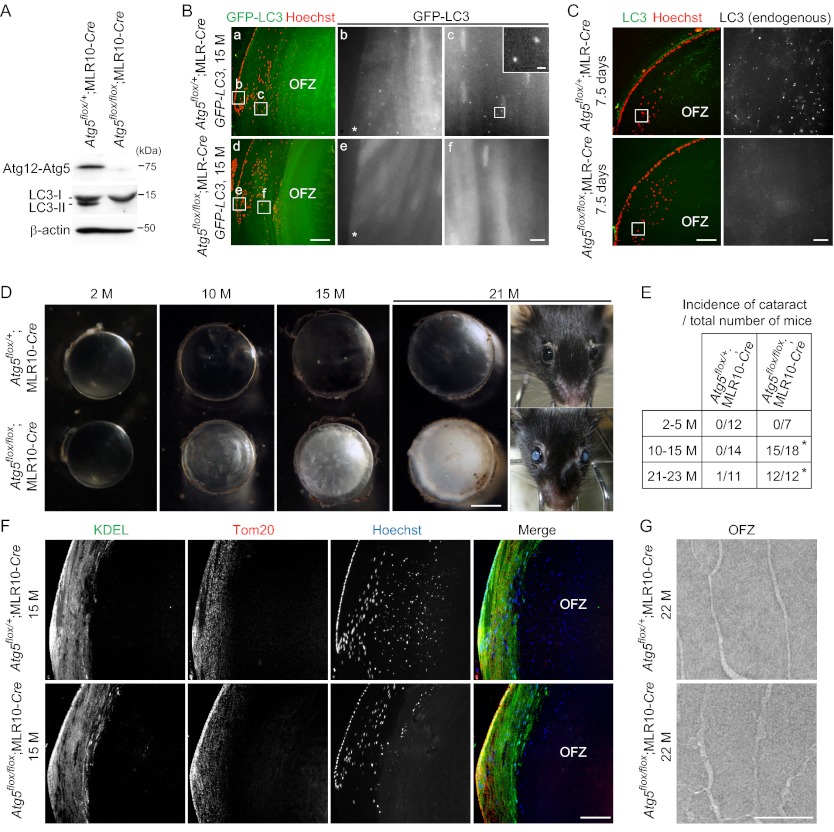
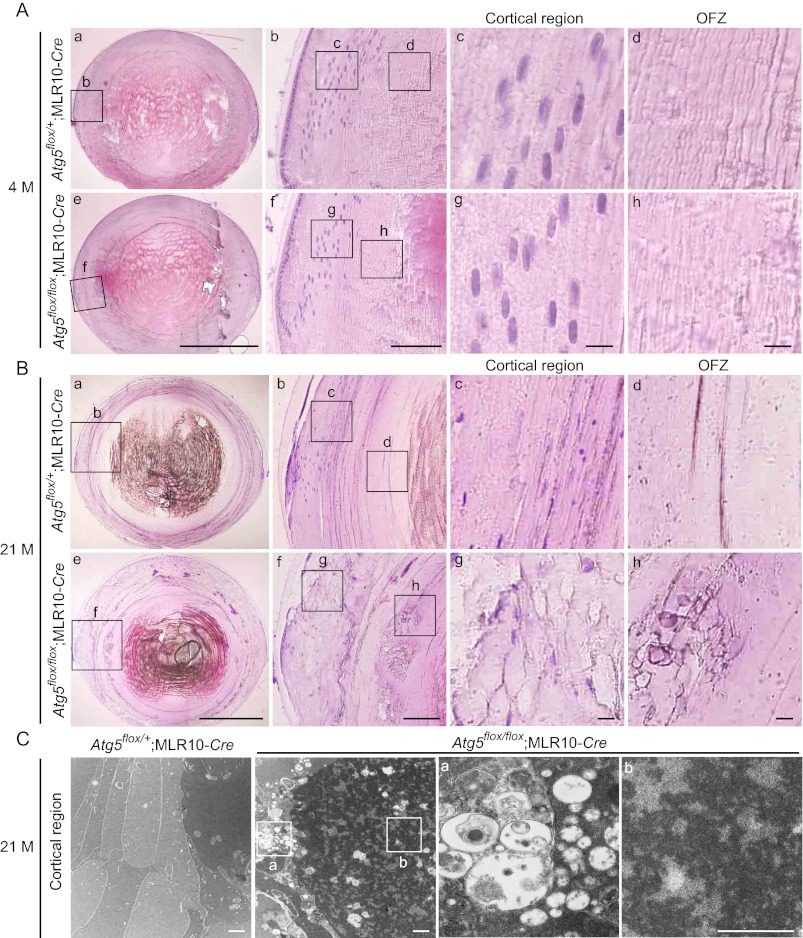
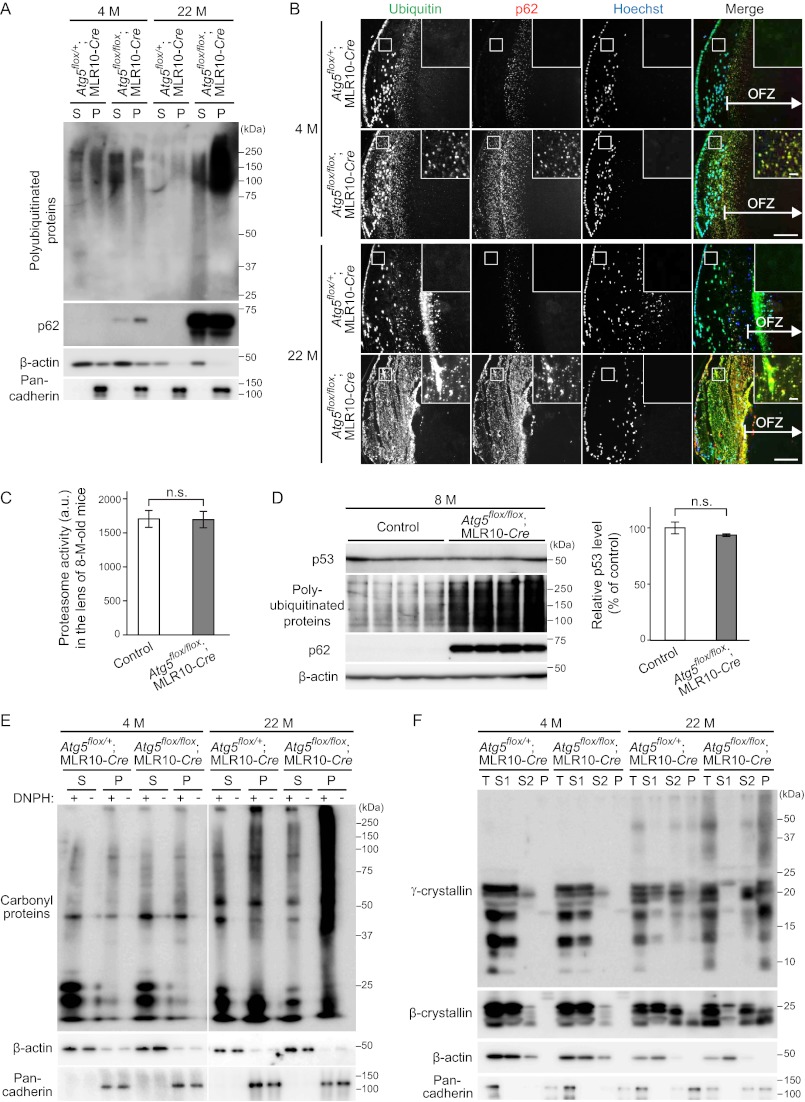
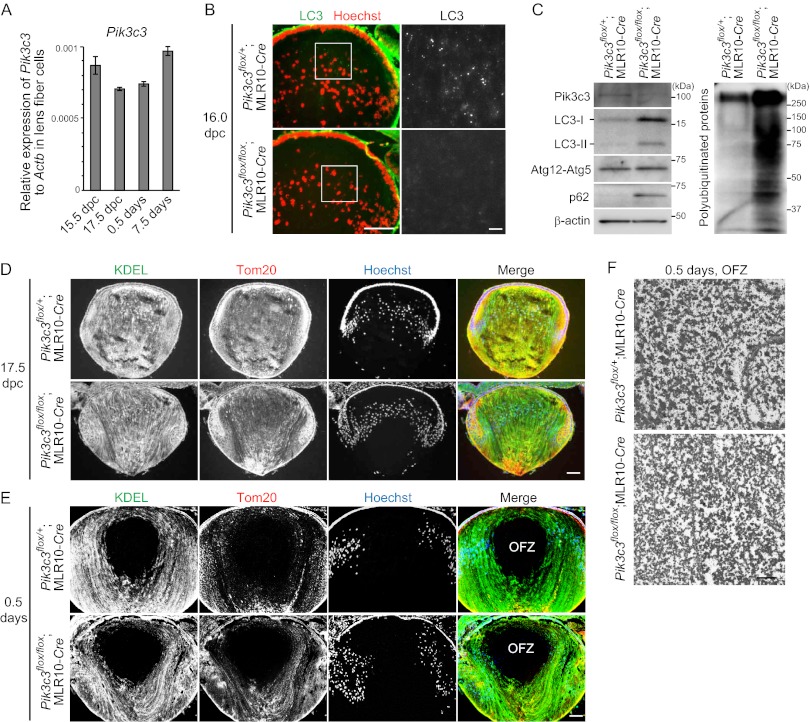
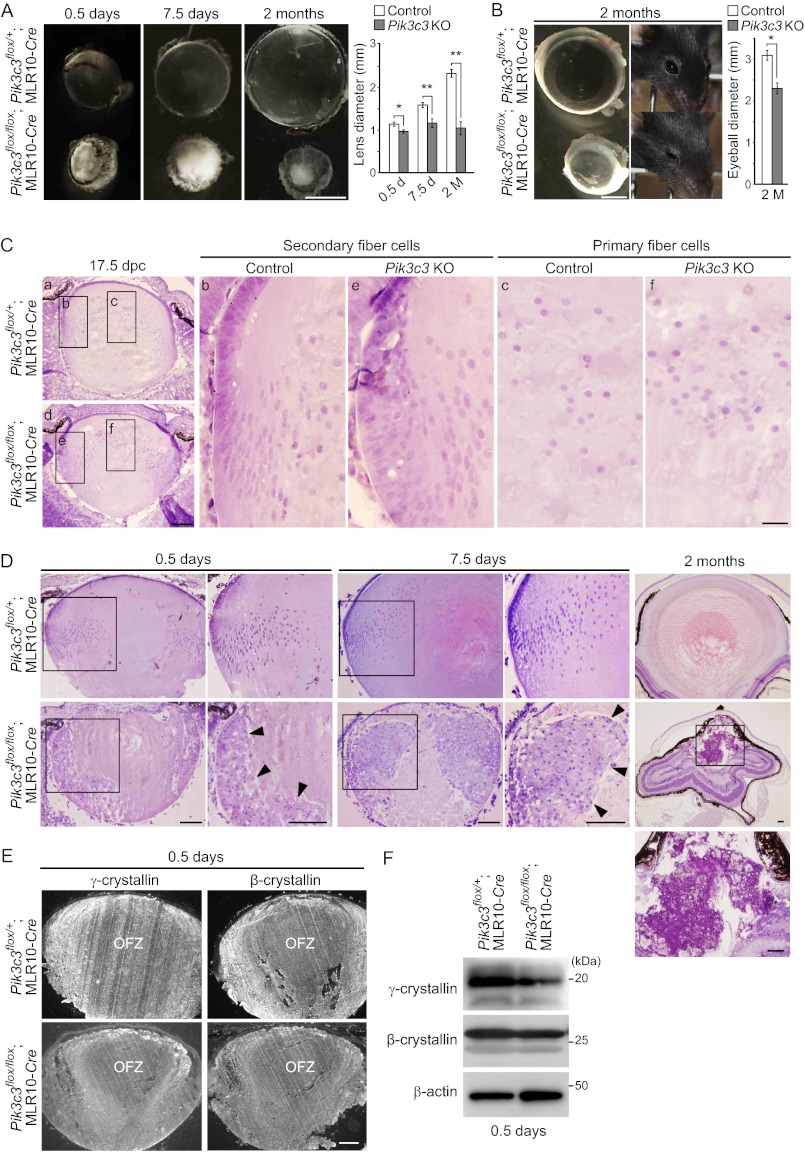
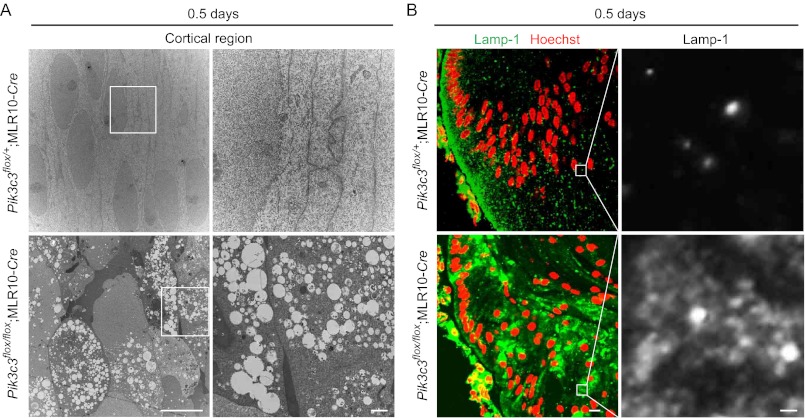
The lens of the eye is composed of fiber cells, which differentiate from epithelial cells and undergo programmed organelle degradation during terminal differentiation. Although autophagy, a major intracellular degradation system, is constitutively active in these cells, its physiological role has remained unclear. We have previously shown that Atg5-dependent macroautophagy is not necessary for lens organelle degradation, at least during the embryonic period. Here, we generated lens-specific Atg5 knock-out mice and showed that Atg5 is not required for lens organelle degradation at any period of life. However, deletion of Atg5 in the lens results in age-related cataract, which is accompanied by accumulation of polyubiquitinated and oxidized proteins, p62, and insoluble crystallins, suggesting a defect in intracellular quality control. We also produced lens-specific Pik3c3 knock-out mice to elucidate the possible involvement of Atg5-independent alternative autophagy, which is proposed to be dependent on Pik3c3 (also known as Vps34), in lens organelle degradation. Deletion of Pik3c3 in the lens does not affect lens organelle degradation, but it leads to congenital cataract and a defect in lens development after birth likely due to an impairment of the endocytic pathway. Taken together, these results suggest that clearance of lens organelles is independent of macroautophagy. These findings also clarify the physiological role of Atg5 and Pik3c3 in quality control and development of the lens, respectively.
Figures
References
-
- Piatigorsky J. (1981) Lens differentiation in vertebrates. A review of cellular and molecular features. Differentiation 19, 134–153 - PubMed
-
- McAvoy J. W., Chamberlain C. G., de Iongh R. U., Hales A. M., Lovicu F. J. (1999) Lens development. Eye 13, 425–437 - PubMed
-
- Kuwabara T., Imaizumi M. (1974) Denucleation process of the lens. Invest. Ophthalmol. 13, 973–981 - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Medical
Molecular Biology Databases
