

Controlled iontophoresis coupled with fast-scan cyclic voltammetry/electrophysiology in awake, freely moving animals
- PMID: 23480099
- PMCID: PMC3656757
- DOI: 10.1021/cn400031v
Controlled iontophoresis coupled with fast-scan cyclic voltammetry/electrophysiology in awake, freely moving animals
Abstract
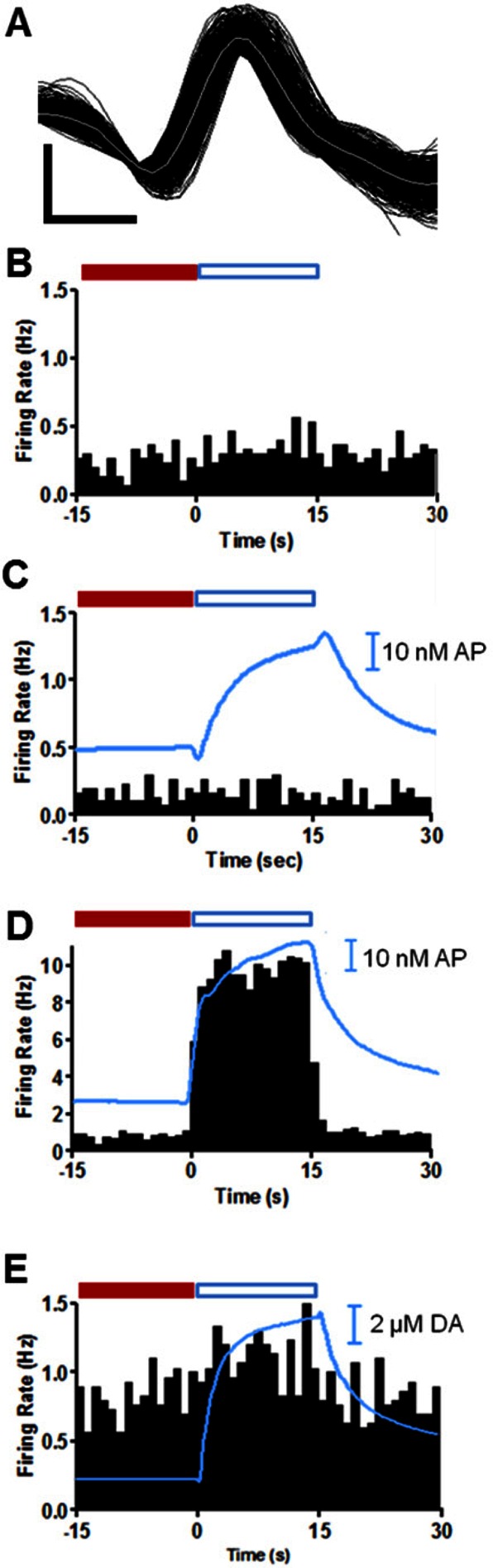
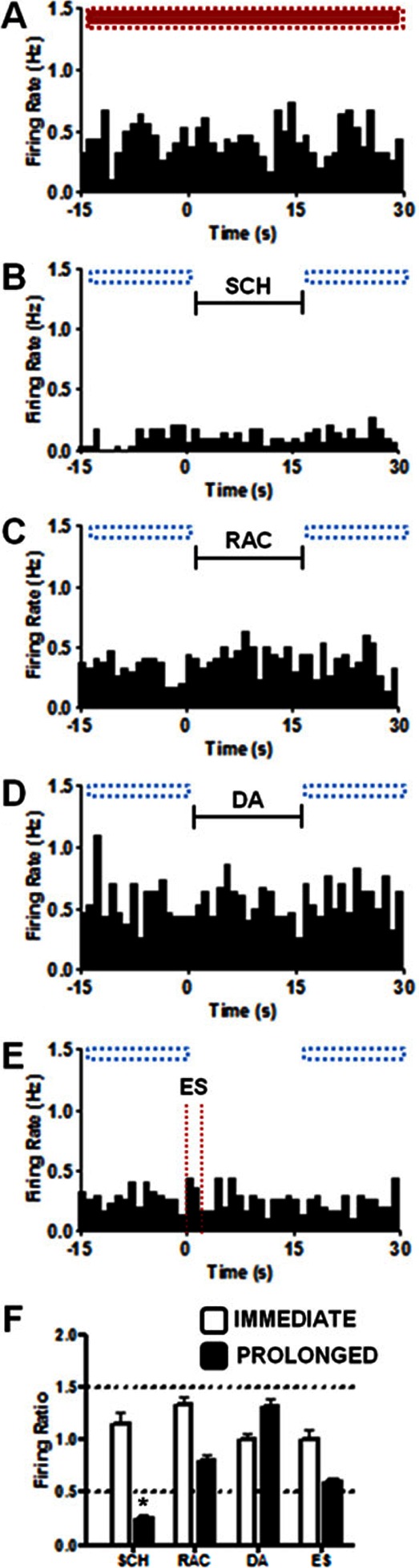
Simultaneous electrochemical and electrophysiological data were recorded to evaluate the effects of controlled local application of dopaminergic agonists and antagonists in awake rats. Measurements were made with a probe consisting of a carbon-fiber microelectrode fused to three iontophoretic barrels used to introduce the drugs of interest. The probe and the manipulator used to position it in the brain of behaving animals were optimized to improve their performance. The effect of the dopamine autoreceptor on electrically stimulated release was demonstrated. Dopamine inhibited the release of endogenous dopamine whereas raclopride, a D2 antagonist, enhanced it, with similar responses in anesthetized and awake animals. We also examined changes in the firing rate of nucleus accumbens (NAc) neurons in awake animals during and after brief (15 s) iontophoretic ejections of SCH 23390 (D1 receptor antagonist) or raclopride. Changes in response to these antagonists were seen both immediately and on a prolonged time scale. Application of raclopride increased the firing rate in 40% of medium spiny neurons (MSNs), of which half responded immediately. Decreases in firing rate were observed in 46% of MSNs after SCH 23390 application. Only 11% of MSNs responded to both antagonists and one MSN (3%) showed no response to either drug. The same prolonged response in firing rate was seen for electrically stimulated and locally applied dopamine in 75% of MSNs. These results are in agreement with previously reported distributions for dopamine receptor subtypes on MSNs and probe the effects of dopamine on these cell populations.
Figures
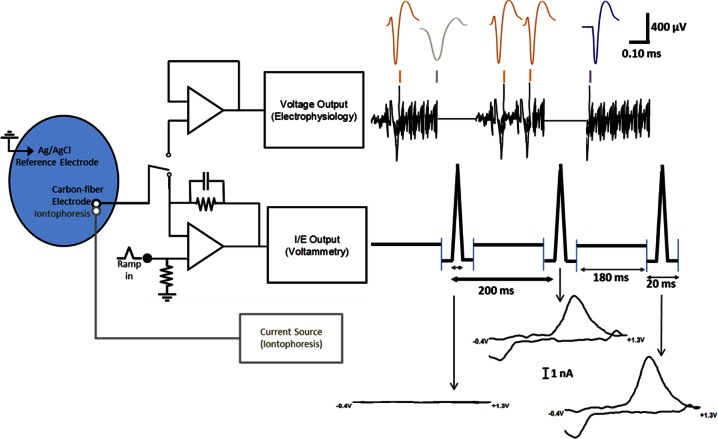
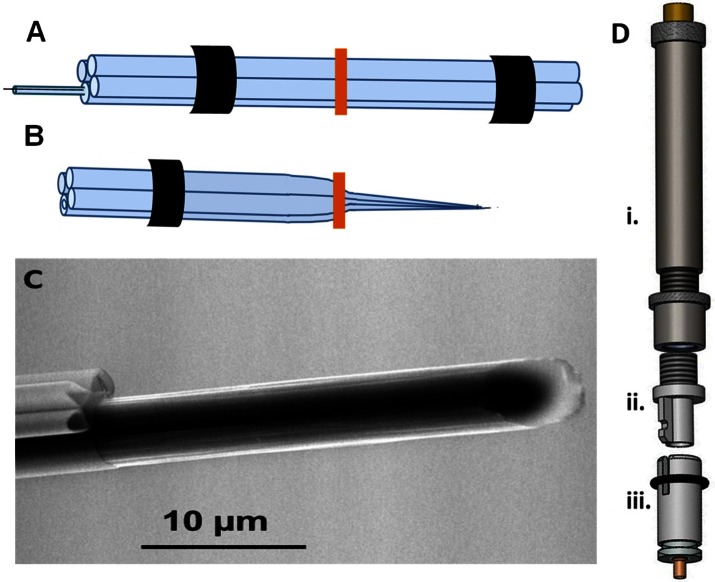
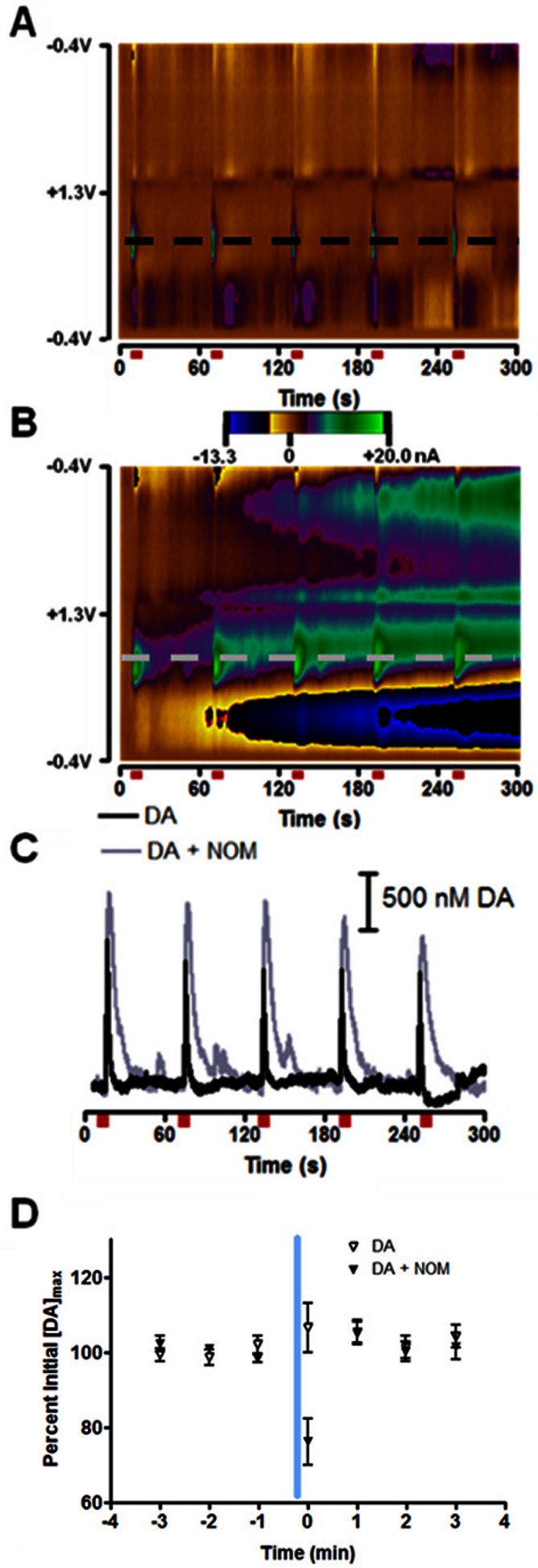
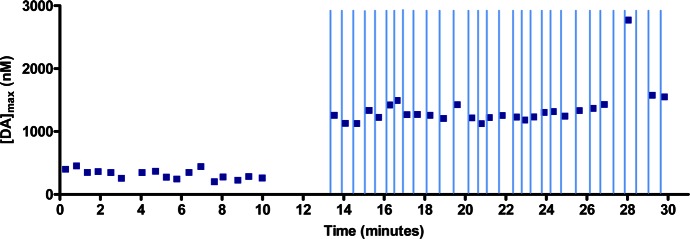
References
-
- Phillips P. E.; Stuber G. D.; Heien M. L.; Wightman R. M.; Carelli R. M. (2003) Subsecond dopamine release promotes cocaine seeking. Nature 422, 614–618. - PubMed
-
- Robinson D. L., and Wightman R. M. (2007) Rapid dopamine release in freely moving rats. In Electrochemical Methods for Neuroscience (Michael A. C., and Borland L. M., Eds.), pp 17–36, CRC Press, Boca Raton, FL. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Research Materials
Miscellaneous
