

Engineering the type III secretion system in non-replicating bacterial minicells for antigen delivery
- PMID: 23481398
- PMCID: PMC3693737
- DOI: 10.1038/ncomms2594
Engineering the type III secretion system in non-replicating bacterial minicells for antigen delivery
Abstract
Type III protein secretion systems are being considered for vaccine development as virtually any protein antigen can be engineered for delivery by these nanomachines into the class I antigen presentation pathway to stimulate antigen-specific CD8(+) T cells. A limitation in the use of this system is that it requires live virulence-attenuated bacteria, which may preclude its use in certain populations such as children and the immunocompromised. Here we report the engineering of the Salmonella Typhimurium type III secretion system in achromosomal, non-replicating nanoparticles derived from bacterial minicells. The engineered system is shown to be functional and capable of delivering heterologous antigens to the class I antigen presentation pathway stimulating immune responses both in vitro and in vivo. This antigen delivery platform offers a novel approach for vaccine development and cellular immunotherapy.
Conflict of interest statement
Figures
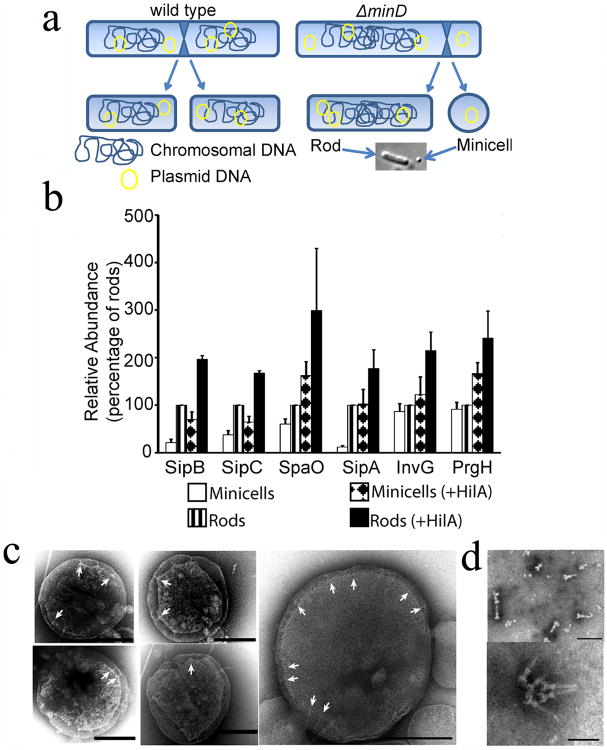
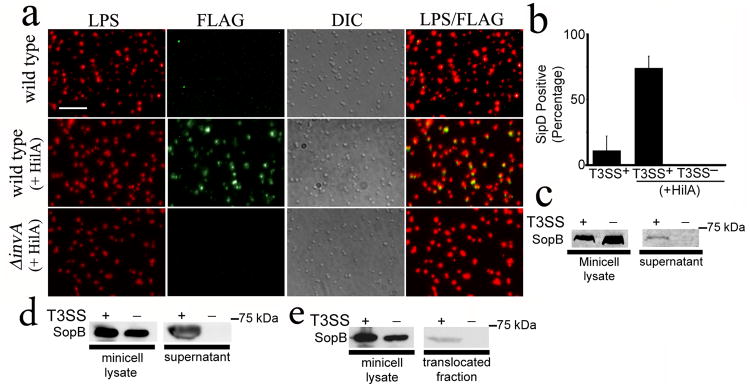
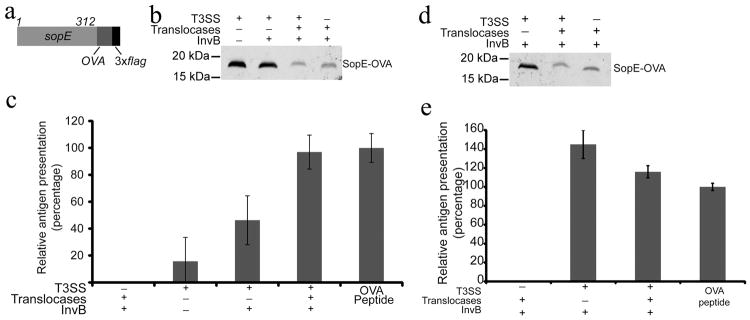
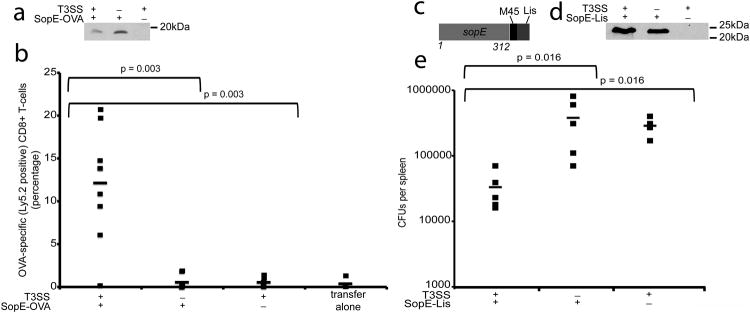
References
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Research Materials
