

Advanced glycation end products acutely impair ca(2+) signaling in bovine aortic endothelial cells
- PMID: 23483845
- PMCID: PMC3593230
- DOI: 10.3389/fphys.2013.00038
Advanced glycation end products acutely impair ca(2+) signaling in bovine aortic endothelial cells
Abstract
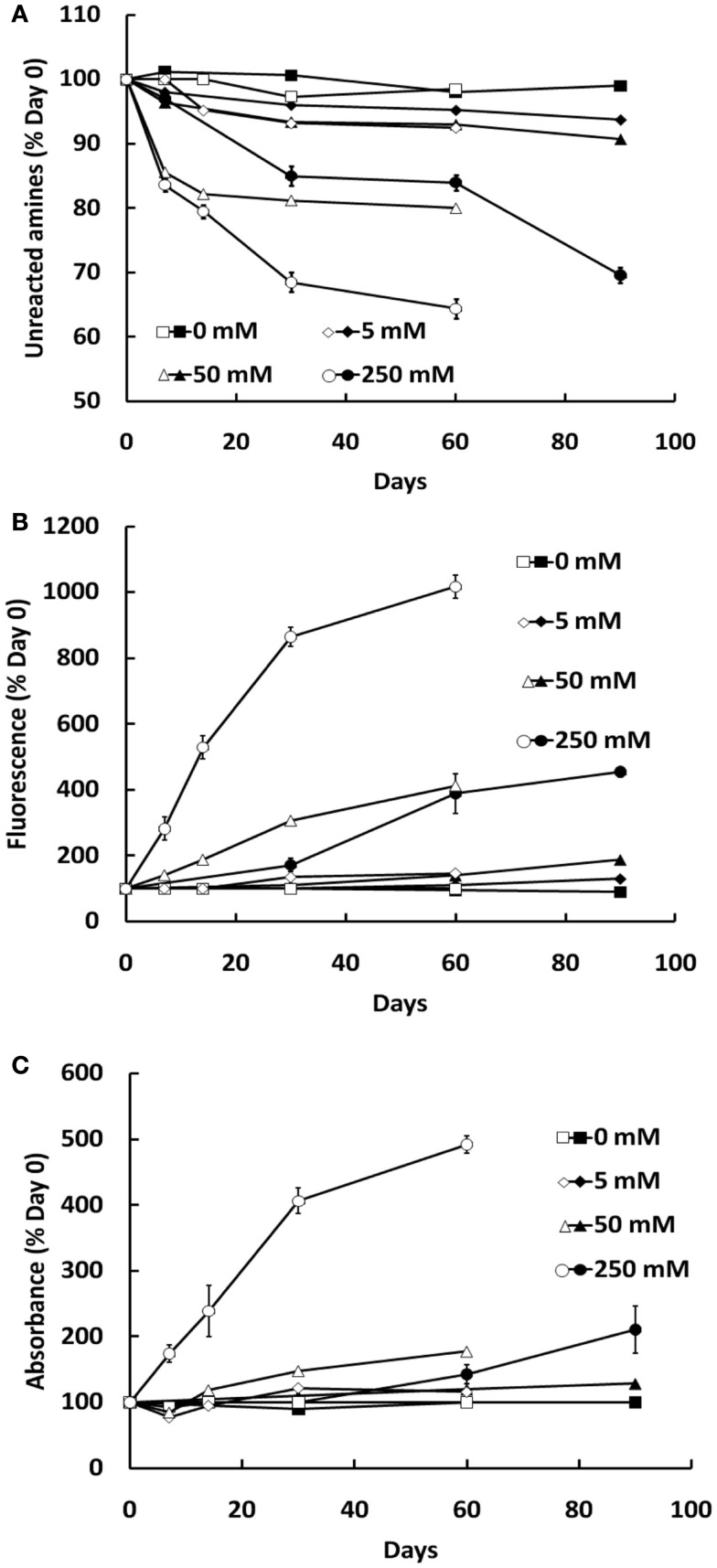
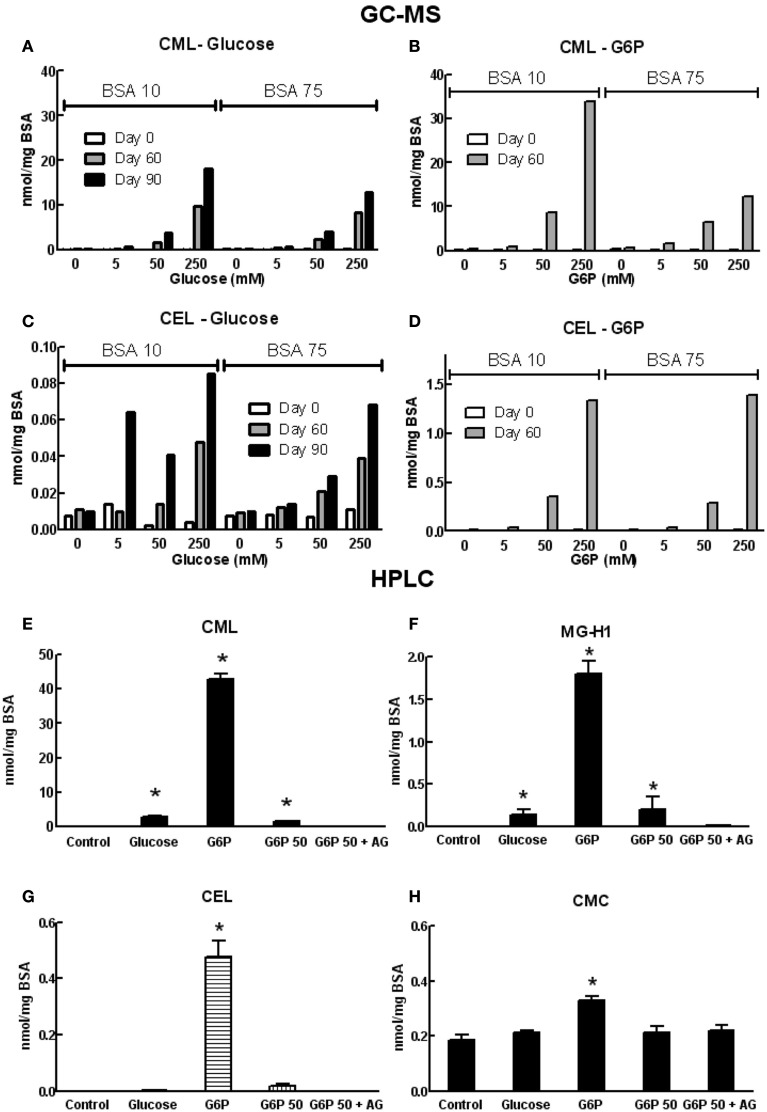
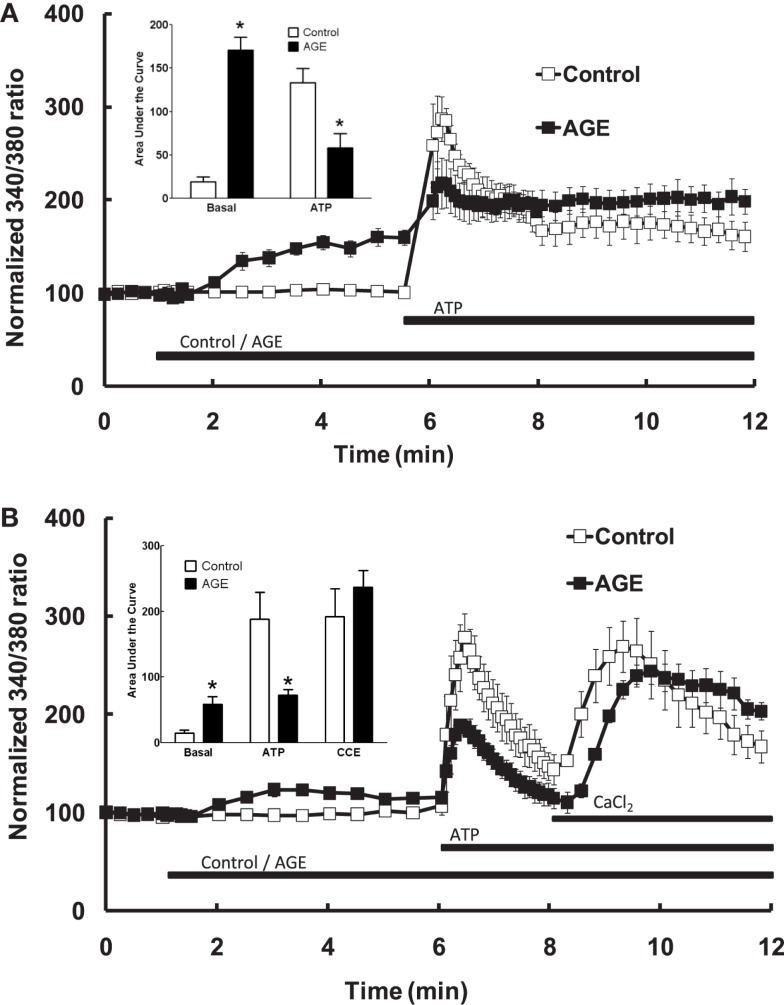
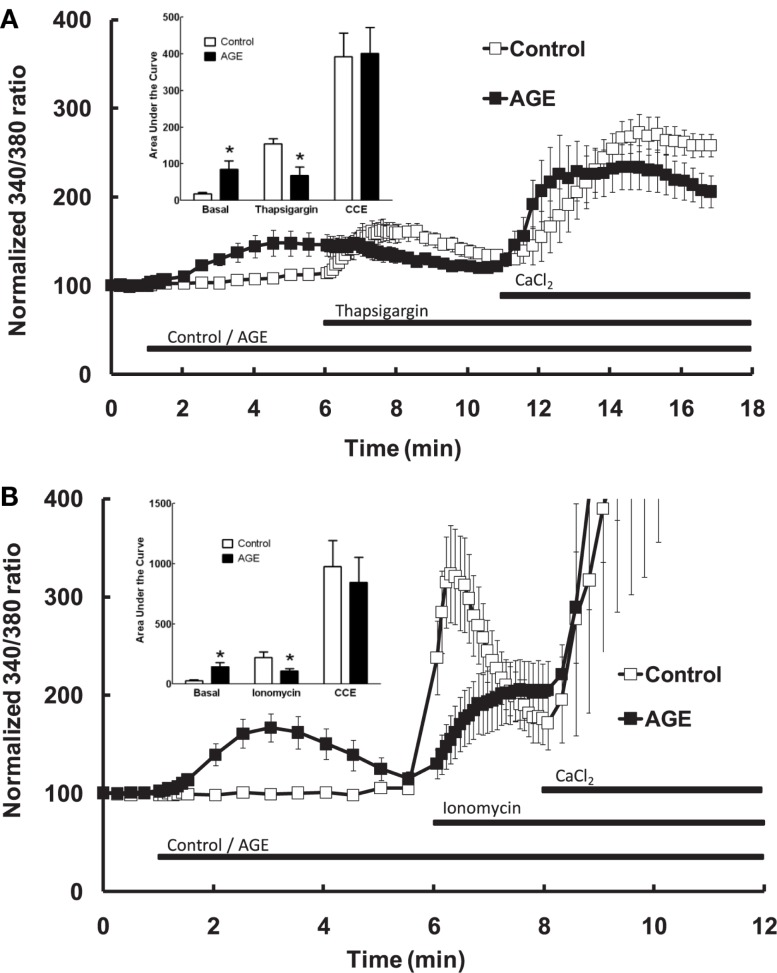
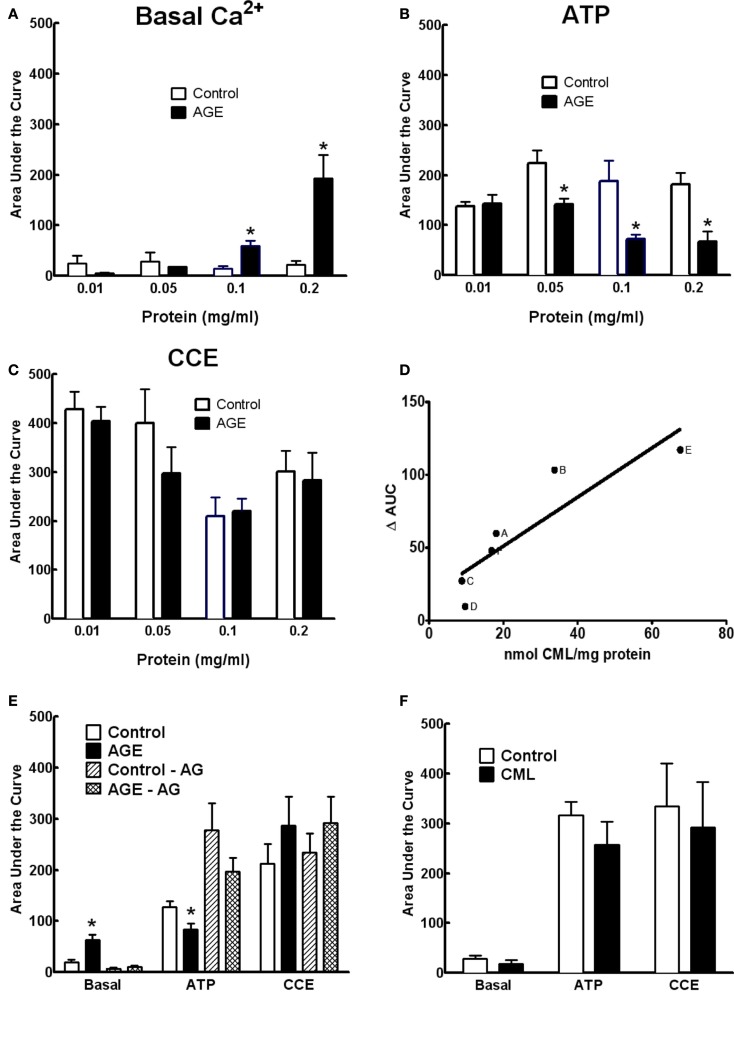
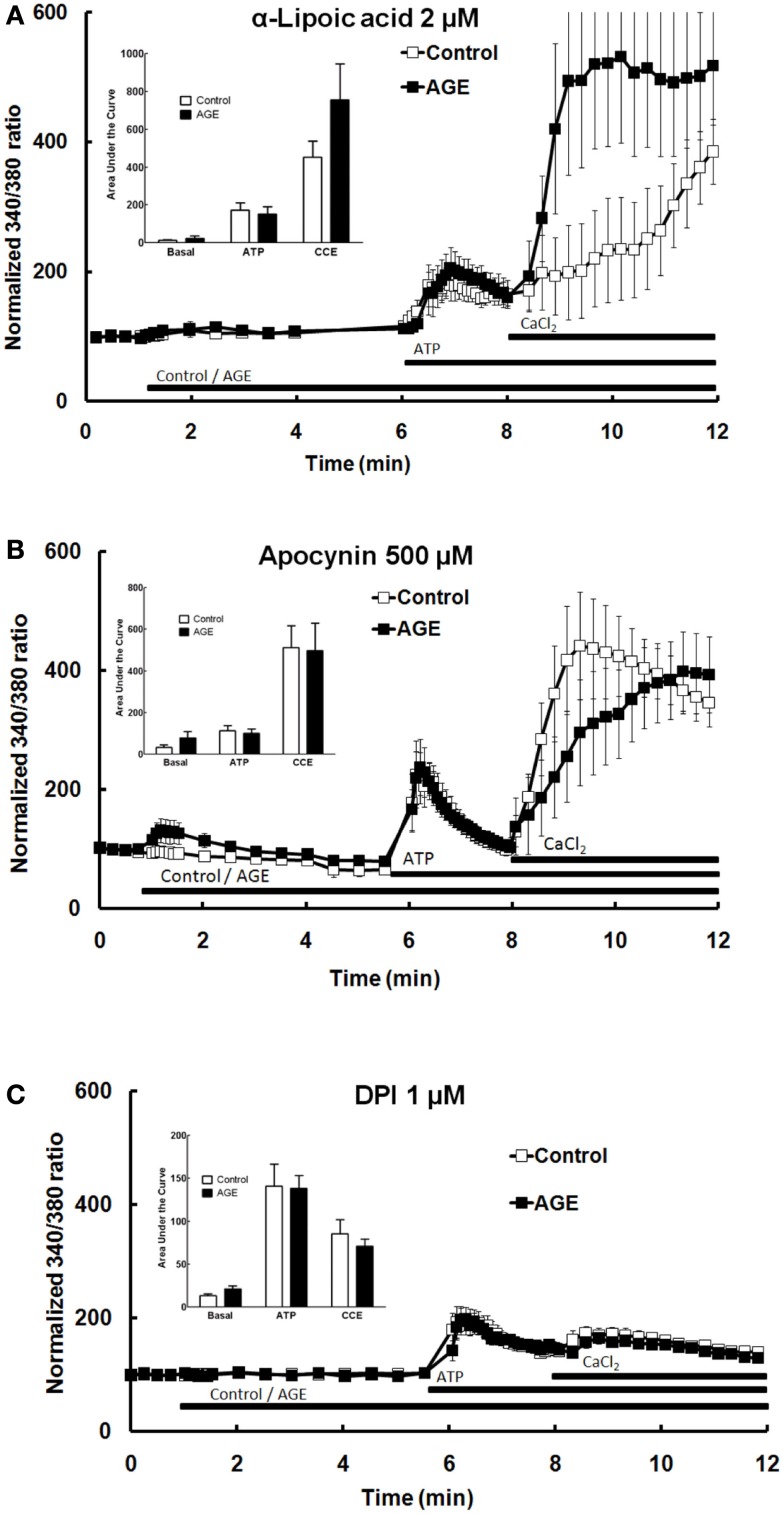
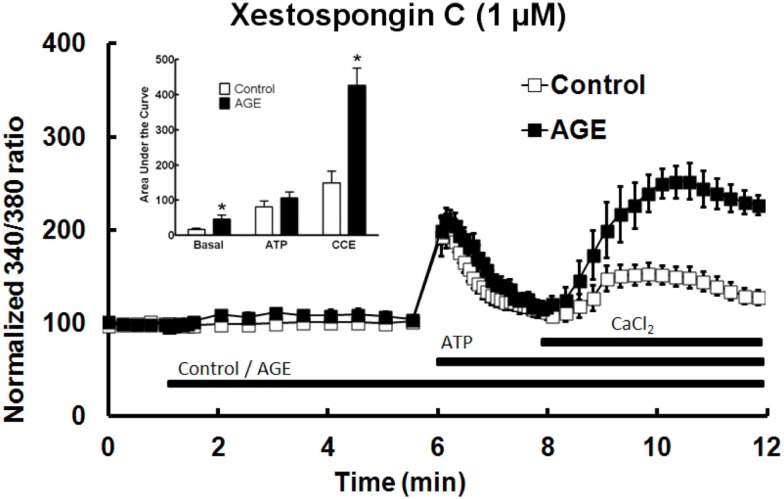
Post-translational modification of proteins in diabetes, including formation of advanced glycation end products (AGEs) are believed to contribute to vascular dysfunction and disease. Impaired function of the endothelium is an early indicator of vascular dysfunction in diabetes and as many endothelial cell processes are dependent upon intracellular [Ca(2+)] and Ca(2+) signaling, the aim of this study was to examine the acute effects of AGEs on Ca(2+) signaling in bovine aortic endothelial cells (BAEC). Ca(2+) signaling was studied using the fluorescent indicator dye Fura-2-AM. AGEs were generated by incubating bovine serum albumin with 0-250 mM glucose or glucose-6-phosphate for 0-120 days at 37°C. Under all conditions, the main AGE species generated was carboxymethyl lysine (CML) as assayed using both gas-liquid chromatograph-mass spectroscopy and high-performance liquid chromatography. In Ca(2+)-replete solution, exposure of BAEC to AGEs for 5 min caused an elevation in basal [Ca(2+)] and attenuated the increase in intracellular [Ca(2+)] caused by ATP (100 μM). In the absence of extracellular Ca(2+), exposure of BAEC to AGEs for 5 min caused an elevation in basal [Ca(2+)] and attenuated subsequent intracellular Ca(2+) release caused by ATP, thapsigargin (0.1 μM), and ionomycin (3 μM), but AGEs did not affect extracellular Ca(2+) entry induced by the re-addition of Ca(2+) to the bathing solution in the presence of any of these agents. The anti-oxidant α-lipoic acid (2 μM) and NAD(P)H oxidase inhibitors apocynin (500 μM) and diphenyleneiodonium (1 μM) abolished these effects of AGEs on BAECs, as did the IP3 receptor antagonist xestospongin C (1 μM). In summary, AGEs caused an acute depletion of Ca(2+) from the intracellular store in BAECs, such that the Ca(2+) signal stimulated by the subsequent application other agents acting upon this store is reduced. The mechanism may involve generation of reactive oxygen species from NAD(P)H oxidase and possible activation of the IP3 receptor.
Keywords: advanced glycation end products; calcium signaling; endothelium; reactive oxygen species.
Figures

References
-
- Alkhalaf A., Kleefstra N., Groenier K. H., Bilo H. J., Gans R. O., Heeringa P., et al. (2012). Effect of benfotiamine on advanced glycation endproducts and markers of endothelial dysfunction and inflammation in diabetic nephropathy. PLoS ONE 7:e40427.10.1371/journal.pone.0040427 - DOI - PMC - PubMed
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Miscellaneous