

Divergent roles for Wnt/β-catenin signaling in epithelial maintenance and breakdown during semicircular canal formation
- PMID: 23487315
- PMCID: PMC3621490
- DOI: 10.1242/dev.092882
Divergent roles for Wnt/β-catenin signaling in epithelial maintenance and breakdown during semicircular canal formation
Abstract
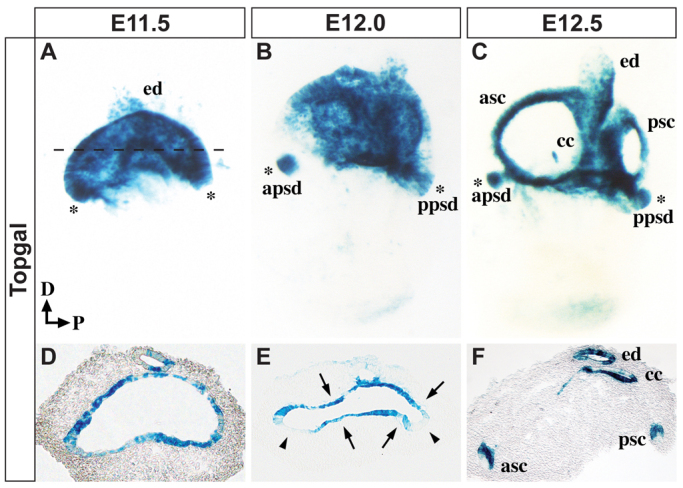
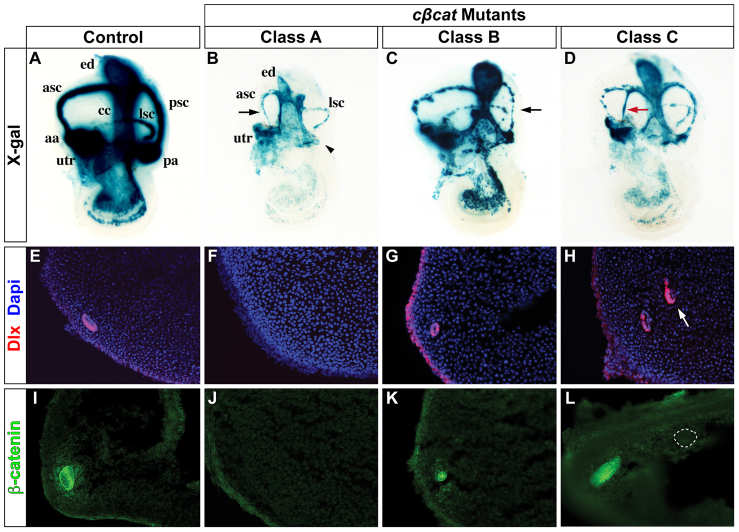
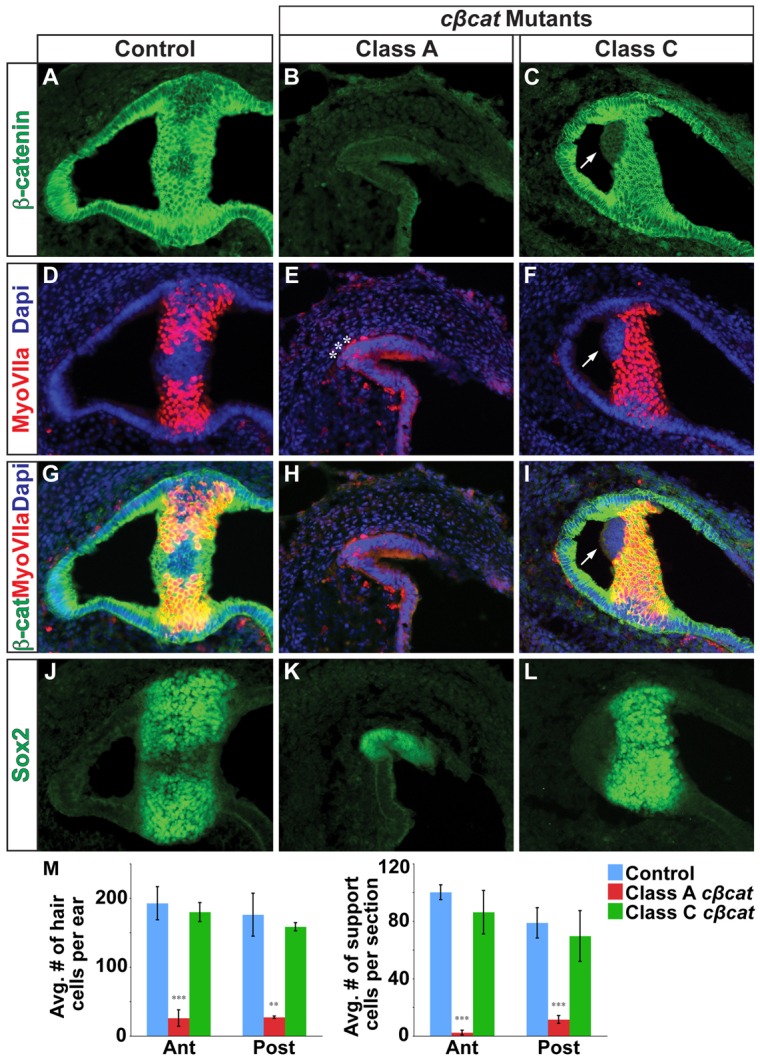
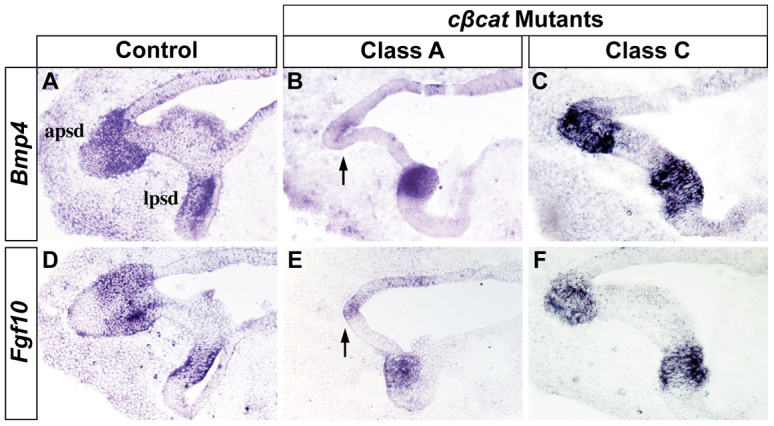
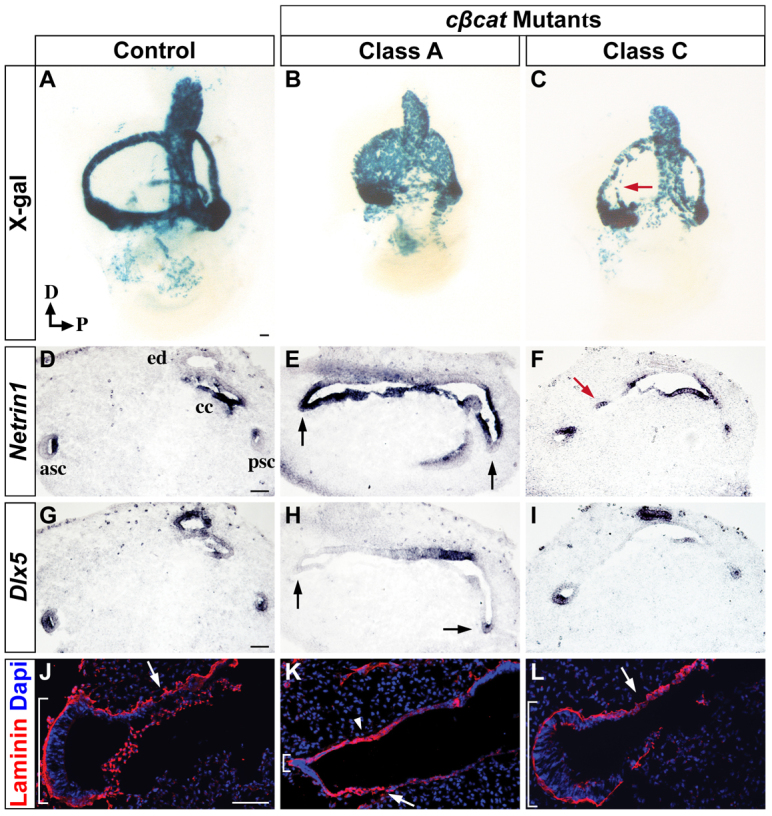
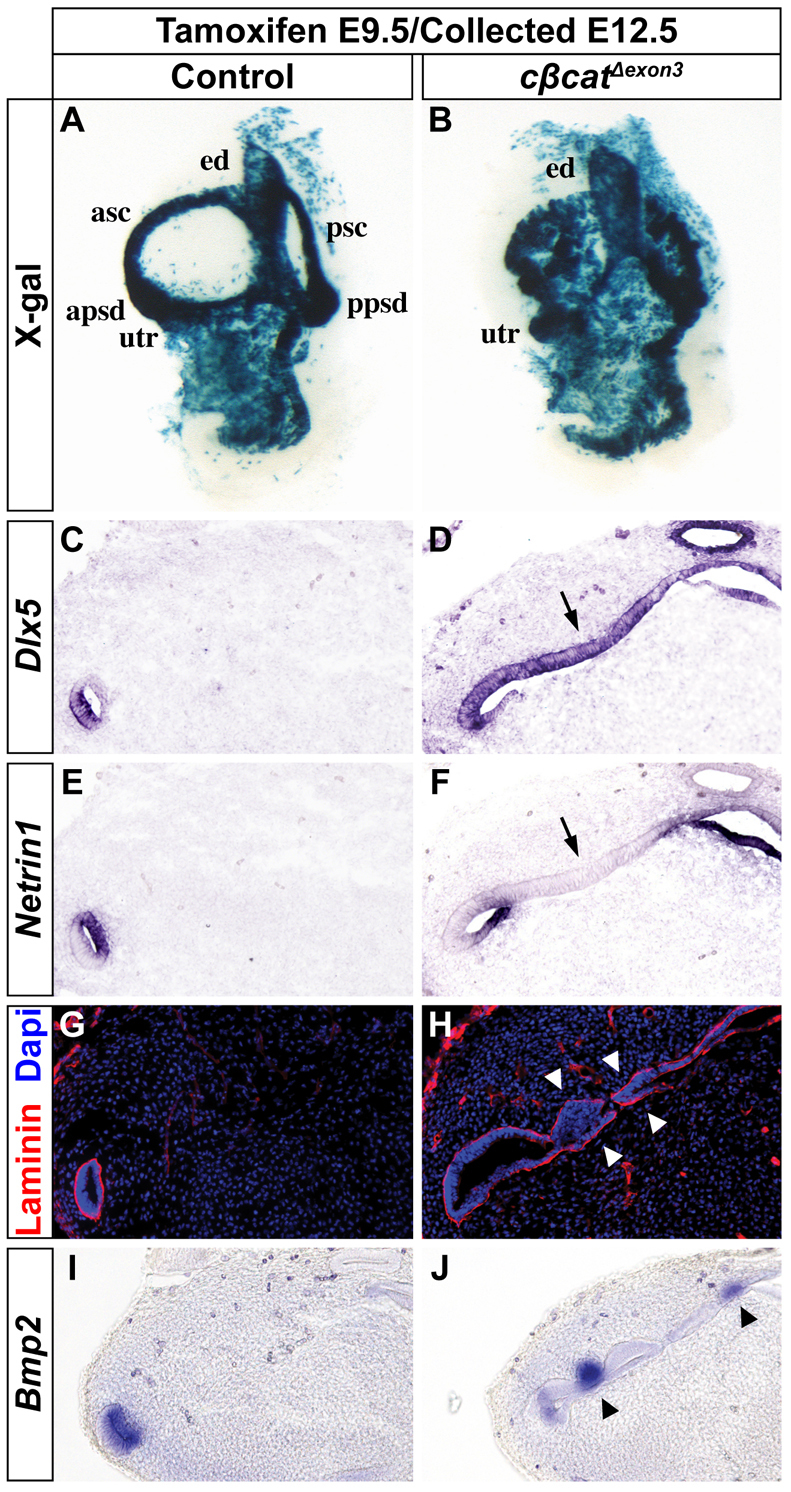
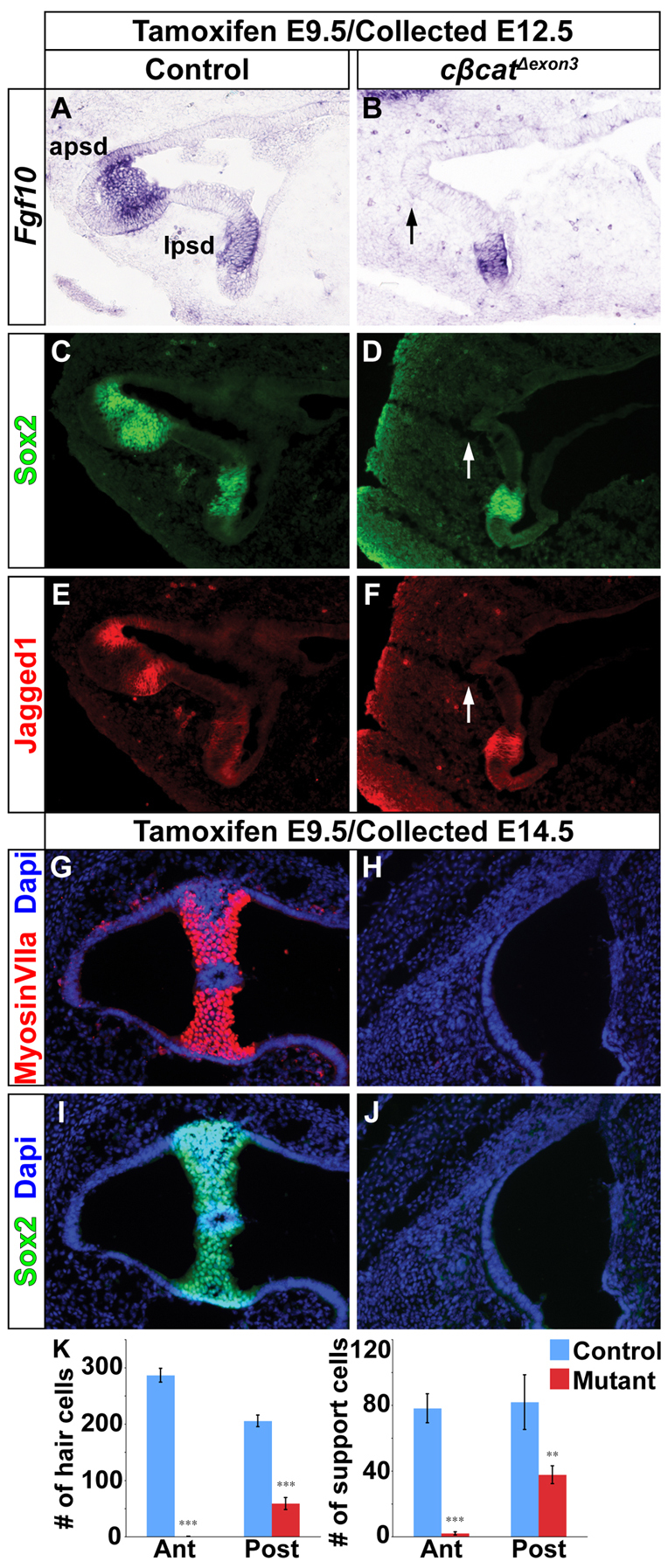
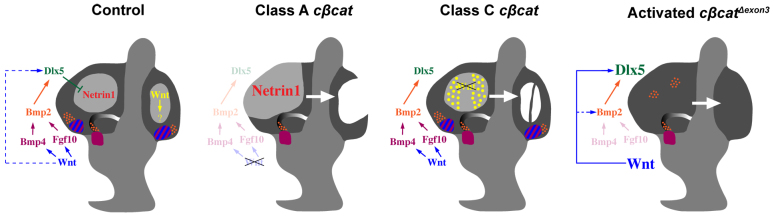
The morphogenetic program that shapes the three semicircular canals (SSCs) must be executed with extreme precision to satisfy their complex vestibular function. The SSCs emerge from epithelial outgrowths of the dorsal otocyst, the central regions of which fuse and resorb to leave three fluid-filled canals. The Wnt/β-catenin signaling pathway is active at multiple stages of otic development, including during vestibular morphogenesis. How Wnt/β-catenin functionally integrates with other signaling pathways to sculpt the SSCs and their sensory patches is unknown. We used a genetic strategy to spatiotemporally modulate canonical Wnt signaling activity during SSC development in mice. Our findings demonstrate that Wnt/β-catenin signaling functions in a multifaceted manner during SSC formation. In the early phase, Wnt/β-catenin signaling is required to preserve the epithelial integrity of the vertical canal pouch perimeter (presumptive anterior and posterior SSCs) by establishing a sensory-dependent signaling relay that maintains expression of Dlx5 and opposes expression of the fusion plate marker netrin 1. Without this Wnt signaling activity the sensory to non-sensory signaling cascade fails to be activated, resulting in loss of vestibular hair and support cells and the anterior and posterior SSCs. In the later phase, Wnt/β-catenin signaling becomes restricted to the fusion plate where it facilitates the timely resorption of this tissue. Mosaic recombination of β-catenin in small clusters of canal pouch cells prevents their resorption, causing instead the formation of ectopic SSCs. Together, these disparate functions of the Wnt/β-catenin pathway in epithelial maintenance and resorption help regulate the size, shape and number of SSCs.
Figures
References
-
- Bok J., Chang W., Wu D. K. (2007). Patterning and morphogenesis of the vertebrate inner ear. Int. J. Dev. Biol. 51, 521–533 - PubMed
-
- Brault V., Moore R., Kutsch S., Ishibashi M., Rowitch D. H., McMahon A. P., Sommer L., Boussadia O., Kemler R. (2001). Inactivation of the beta-catenin gene by Wnt1-Cre-mediated deletion results in dramatic brain malformation and failure of craniofacial development. Development 128, 1253–1264 - PubMed
-
- Chang W., Brigande J. V., Fekete D. M., Wu D. K. (2004). The development of semicircular canals in the inner ear: role of FGFs in sensory cristae. Development 131, 4201–4211 - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
