

Robust inducible Cre recombinase activity in the human malaria parasite Plasmodium falciparum enables efficient gene deletion within a single asexual erythrocytic growth cycle
- PMID: 23489321
- PMCID: PMC3708112
- DOI: 10.1111/mmi.12206
Robust inducible Cre recombinase activity in the human malaria parasite Plasmodium falciparum enables efficient gene deletion within a single asexual erythrocytic growth cycle
Abstract
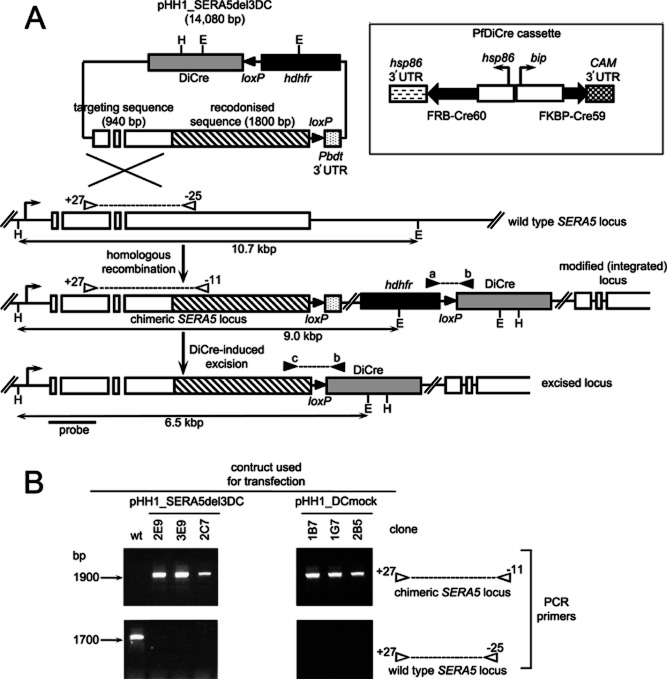
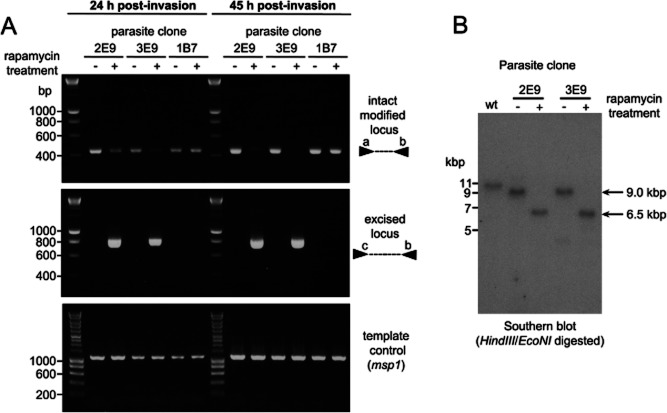
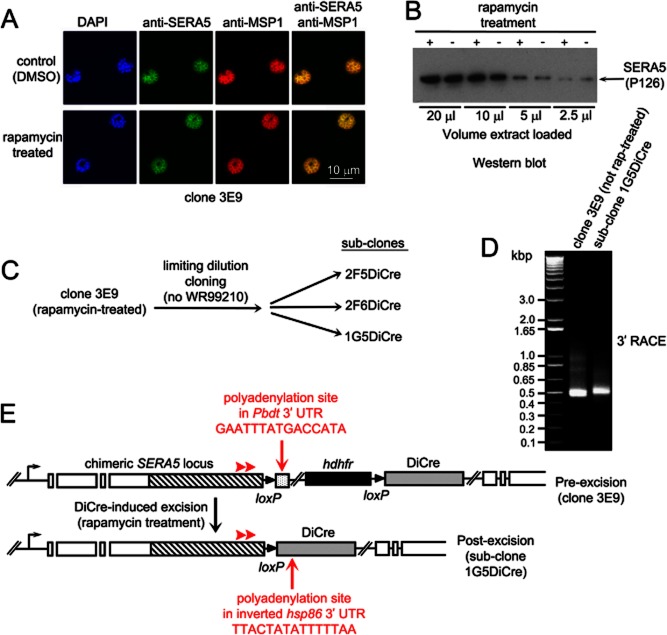
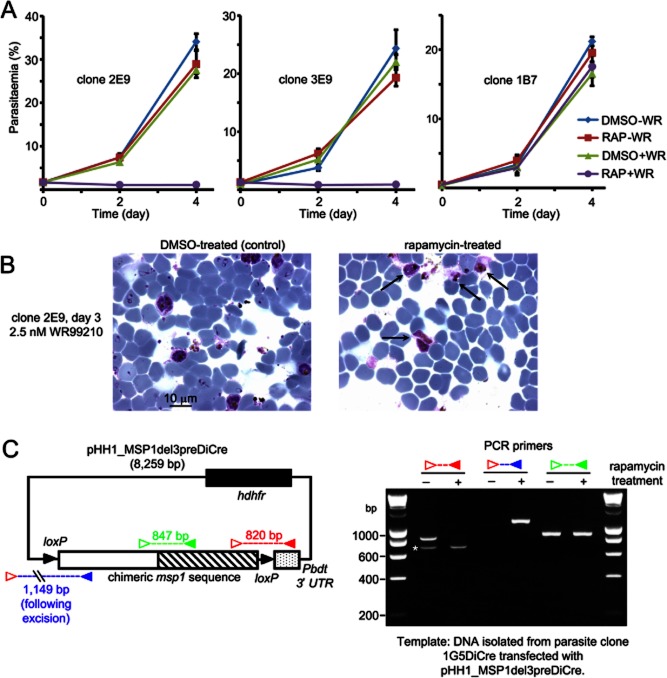
Asexual blood stages of the malaria parasite, which cause all the pathology associated with malaria, can readily be genetically modified by homologous recombination, enabling the functional study of parasite genes that are not essential in this part of the life cycle. However, no widely applicable method for conditional mutagenesis of essential asexual blood-stage malarial genes is available, hindering their functional analysis. We report the application of the DiCre conditional recombinase system to Plasmodium falciparum, the causative agent of the most dangerous form of malaria. We show that DiCre can be used to obtain rapid, highly regulated site-specific recombination in P. falciparum, capable of excising loxP-flanked sequences from a genomic locus with close to 100% efficiency within the time-span of a single erythrocytic growth cycle. DiCre-mediated deletion of the SERA5 3' UTR failed to reduce expression of the gene due to the existence of alternative cryptic polyadenylation sites within the modified locus. However, we successfully used the system to recycle the most widely used drug resistance marker for P. falciparum, human dihydrofolate reductase, in the process producing constitutively DiCre-expressing P. falciparum clones that have broad utility for the functional analysis of essential asexual blood-stage parasite genes.
© 2013 John Wiley & Sons Ltd.
Figures
Comment in
-
From technology to biology: a malaria genetic toolbox for the functional dissection of essential genes.Mol Microbiol. 2013 May;88(4):650-4. doi: 10.1111/mmi.12232. Epub 2013 Apr 24. Mol Microbiol. 2013. PMID: 23614838
Similar articles
-
Generation of a new DiCre expressing parasite strain for functional characterization of Plasmodium falciparum genes in blood stages.Sci Rep. 2024 Oct 15;14(1):24076. doi: 10.1038/s41598-024-75657-x. Sci Rep. 2024. PMID: 39402380 Free PMC article.
-
The dimerisable Cre recombinase allows conditional genome editing in the mosquito stages of Plasmodium berghei.PLoS One. 2020 Oct 12;15(10):e0236616. doi: 10.1371/journal.pone.0236616. eCollection 2020. PLoS One. 2020. PMID: 33044964 Free PMC article.
-
A Novel Tool for the Generation of Conditional Knockouts To Study Gene Function across the Plasmodium falciparum Life Cycle.mBio. 2019 Sep 17;10(5):e01170-19. doi: 10.1128/mBio.01170-19. mBio. 2019. PMID: 31530668 Free PMC article.
-
[In vitro cultivation of Plasmodium falciparum. Applications and limits.- Methodology].Med Trop (Mars). 1982 Jul-Aug;42(4):437-62. Med Trop (Mars). 1982. PMID: 6755144 Review. French.
-
Molecular complexity of sexual development and gene regulation in Plasmodium falciparum.Int J Parasitol. 2004 Dec;34(13-14):1451-8. doi: 10.1016/j.ijpara.2004.10.013. Int J Parasitol. 2004. PMID: 15582522 Review.
Cited by
-
Generation of a new DiCre expressing parasite strain for functional characterization of Plasmodium falciparum genes in blood stages.Sci Rep. 2024 Oct 15;14(1):24076. doi: 10.1038/s41598-024-75657-x. Sci Rep. 2024. PMID: 39402380 Free PMC article.
-
The Plasmodium falciparum rhoptry bulb protein RAMA plays an essential role in rhoptry neck morphogenesis and host red blood cell invasion.PLoS Pathog. 2019 Sep 6;15(9):e1008049. doi: 10.1371/journal.ppat.1008049. eCollection 2019 Sep. PLoS Pathog. 2019. PMID: 31491036 Free PMC article.
-
An Uninvited Seat at the Dinner Table: How Apicomplexan Parasites Scavenge Nutrients from the Host.Microorganisms. 2021 Dec 15;9(12):2592. doi: 10.3390/microorganisms9122592. Microorganisms. 2021. PMID: 34946193 Free PMC article. Review.
-
Optimization of CRISPR/Cas System for Improving Genome Editing Efficiency in Plasmodium falciparum.Front Microbiol. 2021 Jan 8;11:625862. doi: 10.3389/fmicb.2020.625862. eCollection 2020. Front Microbiol. 2021. PMID: 33488567 Free PMC article.
-
Plasmodium blood stage development requires the chromatin remodeller Snf2L.Nature. 2025 Mar;639(8056):1069-1075. doi: 10.1038/s41586-025-08595-x. Epub 2025 Feb 19. Nature. 2025. PMID: 39972139 Free PMC article.
References
-
- Agop-Nersesian C, Pfahler J, Lanzer M, Meissner M. Functional expression of ribozymes in Apicomplexa: towards exogenous control of gene expression by inducible RNA-cleavage. Int J Parasitol. 2008;38:673–681. - PubMed
-
- Armstrong CM, Goldberg DE. An FKBP destabilization domain modulates protein levels in Plasmodium falciparum. Nat Methods. 2007;4:1007–1009. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
