

The disturbance of hippocampal CaMKII/PKA/PKC phosphorylation in early experimental diabetes mellitus
- PMID: 23490331
- PMCID: PMC6493428
- DOI: 10.1111/cns.12084
The disturbance of hippocampal CaMKII/PKA/PKC phosphorylation in early experimental diabetes mellitus
Abstract
Background: Defining the impact of diabetes and related risk factors on brain cognitive function is critically important for patients with diabetes.
Aims: To investigate the alterations in hippocampal serine/threonine kinases signaling in the early phase of type 1 and type 2 diabetic rats.
Methods: Early experimental diabetes mellitus was induced in rats with streptozotocin or streptozotocin/high fat. Changes in the phosphorylation of proteins were determined by immunoblotting and immunohistochemistry.
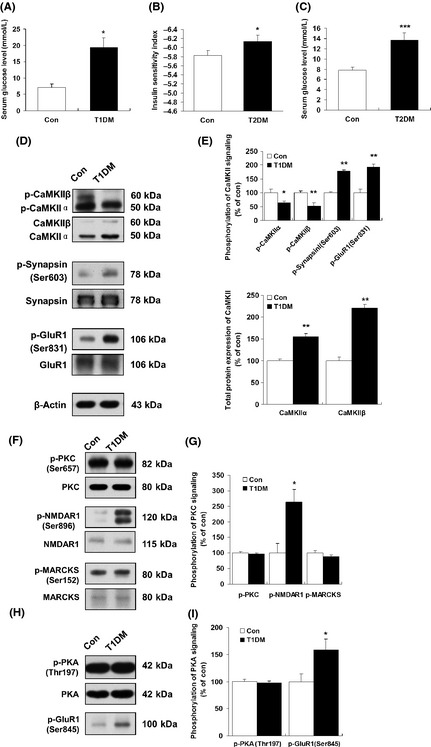
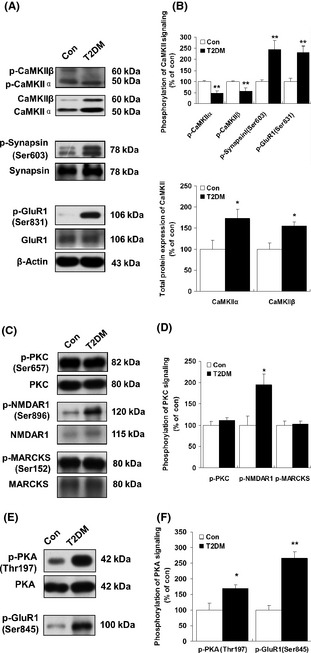
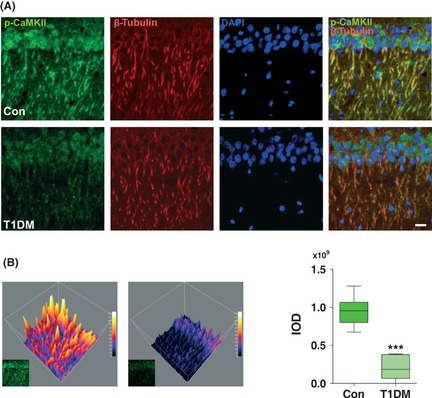
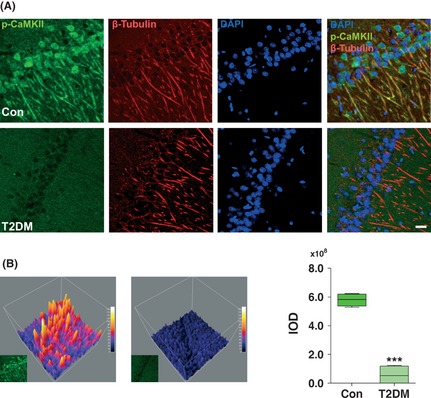
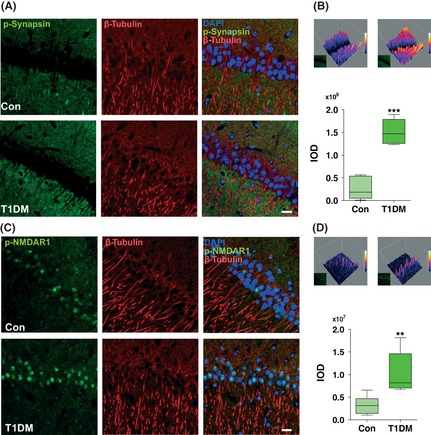
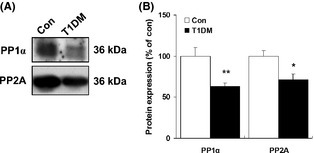
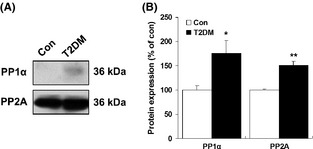
Results: Our data showed a pronounced decrease in the phosphorylation of Ca(2+) /calmodulin-dependent protein kinase II (CaMKII) in the hippocampi of both type 1 and type 2 diabetic rats compared with age-matched control rats. Unexpectedly, we found a significant increase in the phosphorylation of synapsin I (Ser 603) and GluR1 (Ser 831) in the same experiment. In addition, aberrant changes in hippocampal protein kinase C (PKC) and protein kinase A (PKA) signaling in type 1 and type 2 diabetic rats were also found. Moreover, PP1α and PP2A protein levels were decreased in the hippocampus of type 1 diabetic rats, but significantly up-regulated in type 2 diabetic rats.
Conclusions: The disturbance of CaMKII/PKA/PKC phosphorylation in the hippocampus is an early change that may be associated with the development and progression of diabetes-related cognitive dysfunction.
© 2013 Blackwell Publishing Ltd.
Conflict of interest statement
The authors declare no conflict of interest.
Figures
References
-
- Anan F, Masaki T, Shimomura T, et al. Abdominal visceral fat accumulation is associated with hippocampus volume in non‐dementia patients with type 2 diabetes mellitus. Neuroimage 2010;49:57–62. - PubMed
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
Miscellaneous
