

Teaching basic neurophysiology using intact earthworms
- PMID: 23494516
- PMCID: PMC3597421
Teaching basic neurophysiology using intact earthworms
Abstract
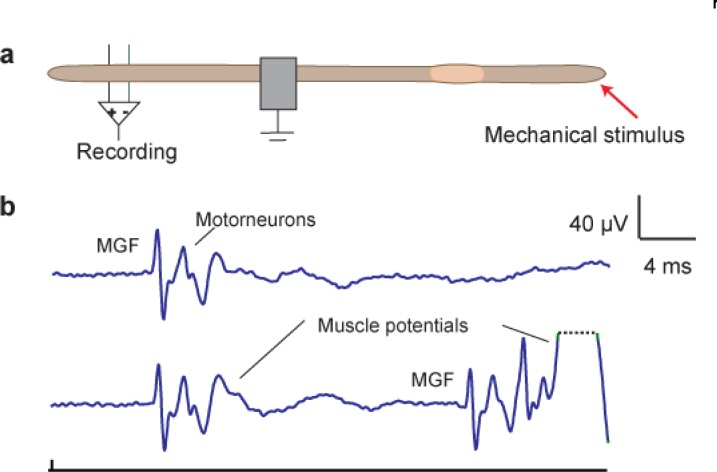
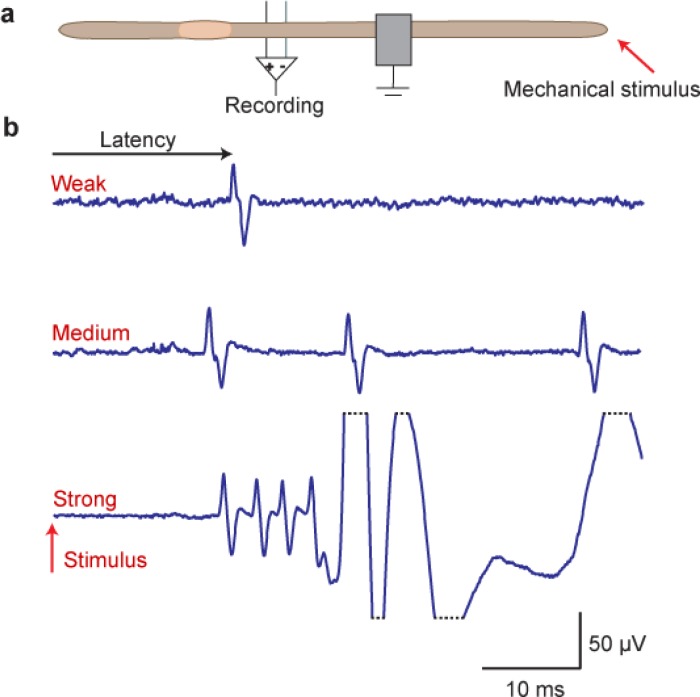
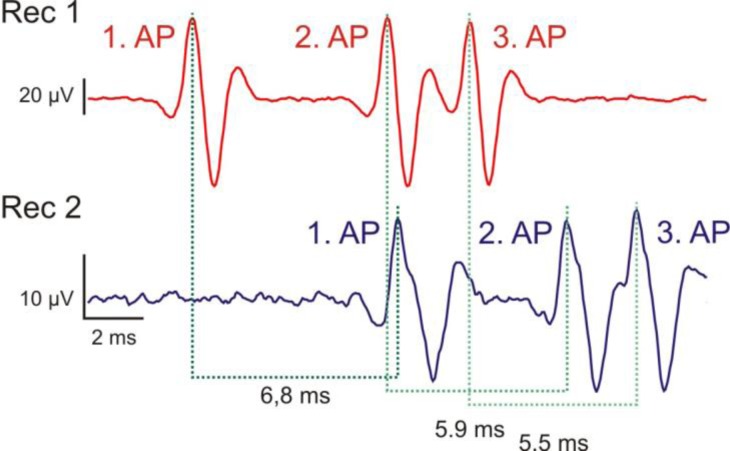
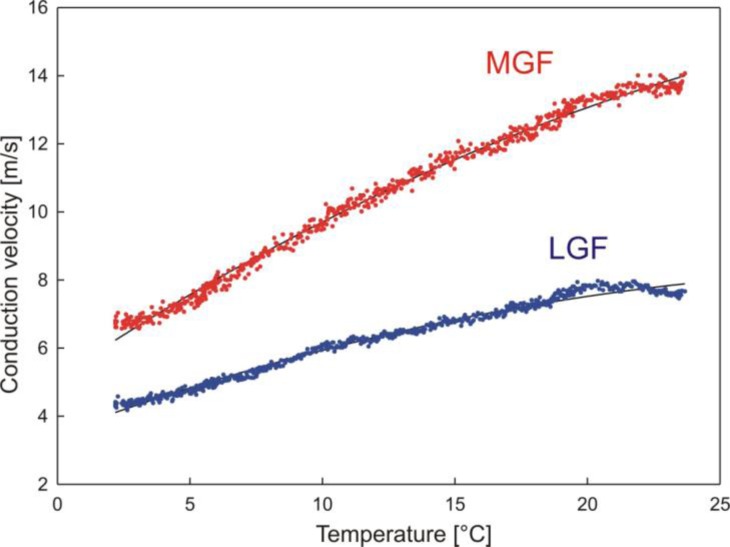
Introductory neurobiology courses face the problem that practical exercises often require expensive equipment, dissections, and a favorable student-instructor ratio. Furthermore, the duration of an experiment might exceed available time or the level of required expertise is too high to successfully complete the experiment. As a result, neurobiological experiments are commonly replaced by models and simulations, or provide only very basic experiments, such as the frog sciatic nerve preparation, which are often time consuming and tedious. Action potential recordings in giant fibers of intact earthworms (Lumbricus terrestris) circumvent many of these problems and result in a nearly 100% success rate. Originally, these experiments were introduced as classroom exercises by Charles Drewes in 1978 using awake, moving earthworms. In 1990, Hans-Georg Heinzel described further experiments using anesthetized earthworms. In this article, we focus on the application of these experiments as teaching tools for basic neurobiology courses. We describe and extend selected experiments, focusing on specific neurobiological principles with experimental protocols optimized for classroom application. Furthermore, we discuss our experience using these experiments in animal physiology and various neurobiology courses at the University of Bonn.
Keywords: action potentials; conduction velocity; earthworms; extracellular recordings; facilitation of conduction velocity; flight reflexes; giant fibers; giant motorneuron; refractory period; spatial size of action potentials; synaptic depression and facilitation; threshold.
Figures
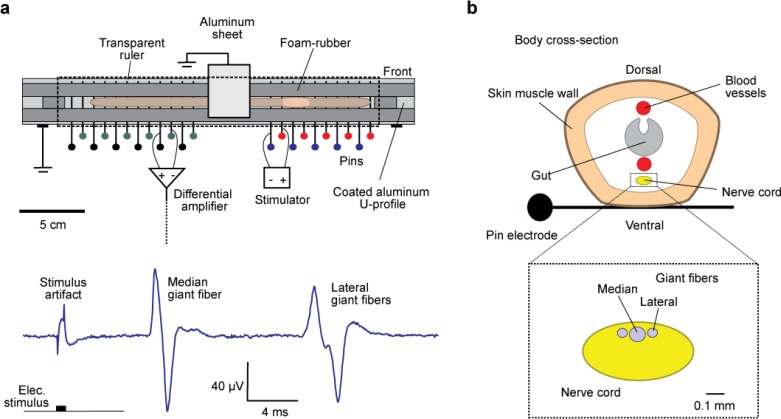
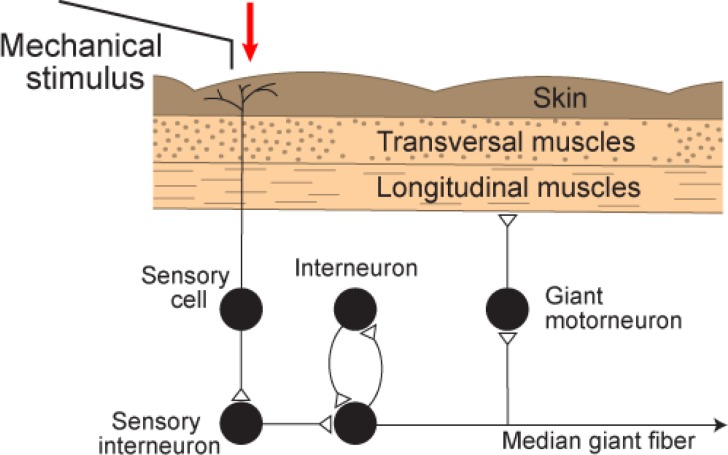
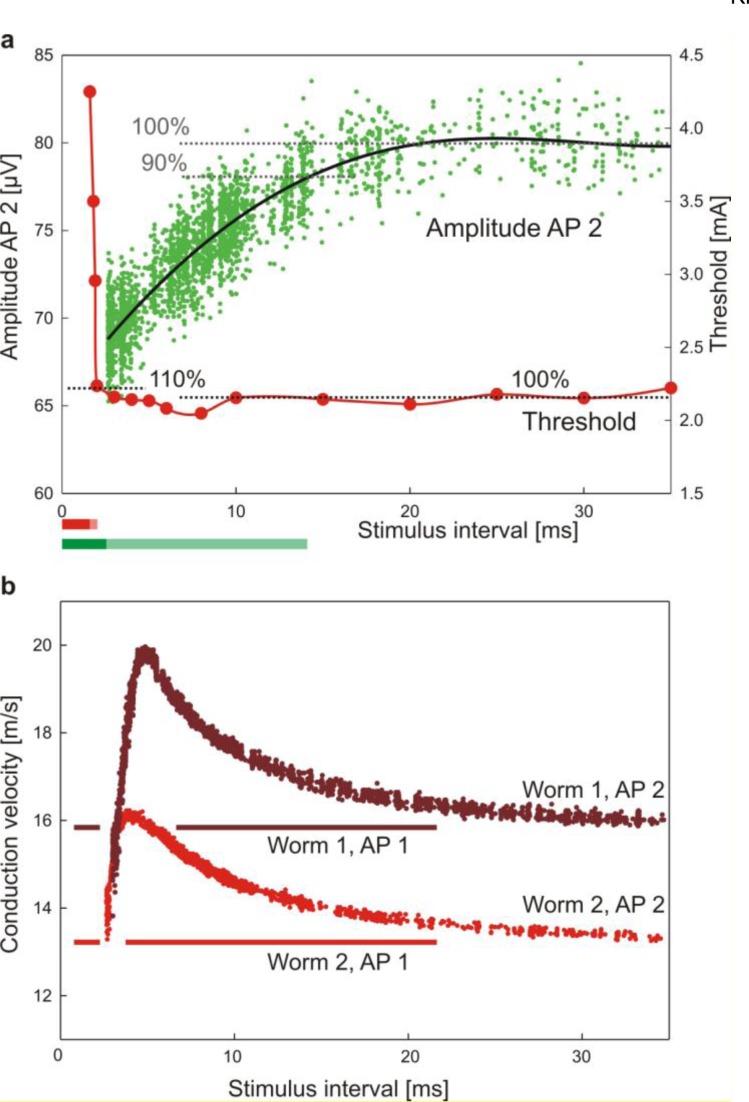
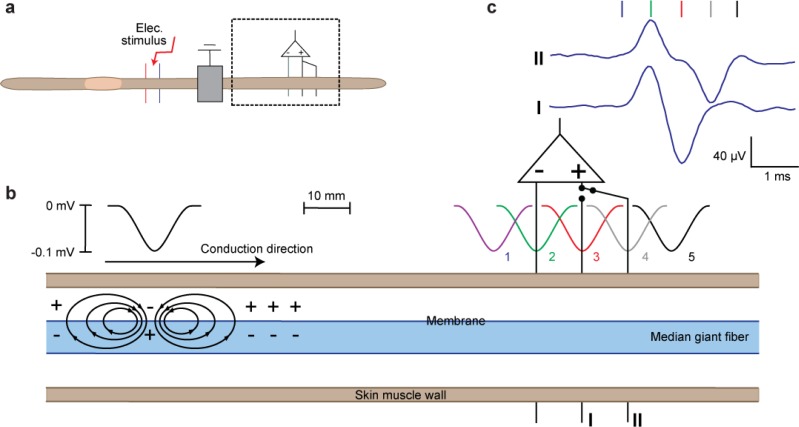
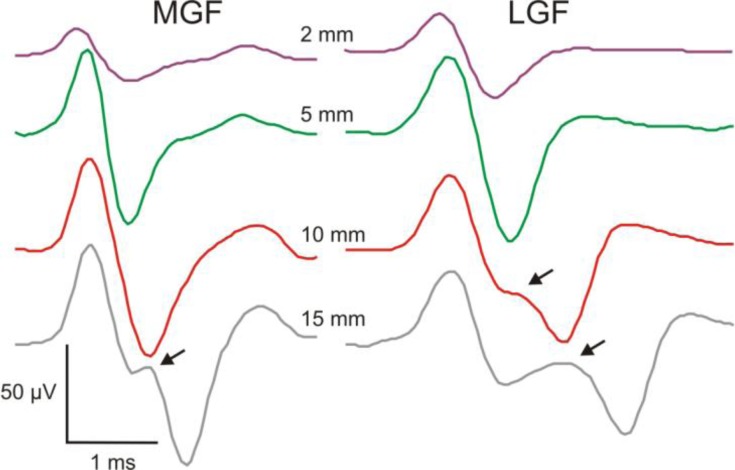
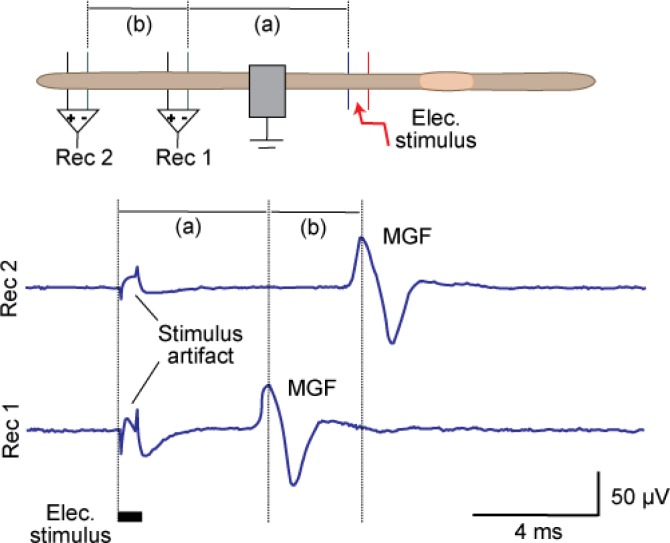
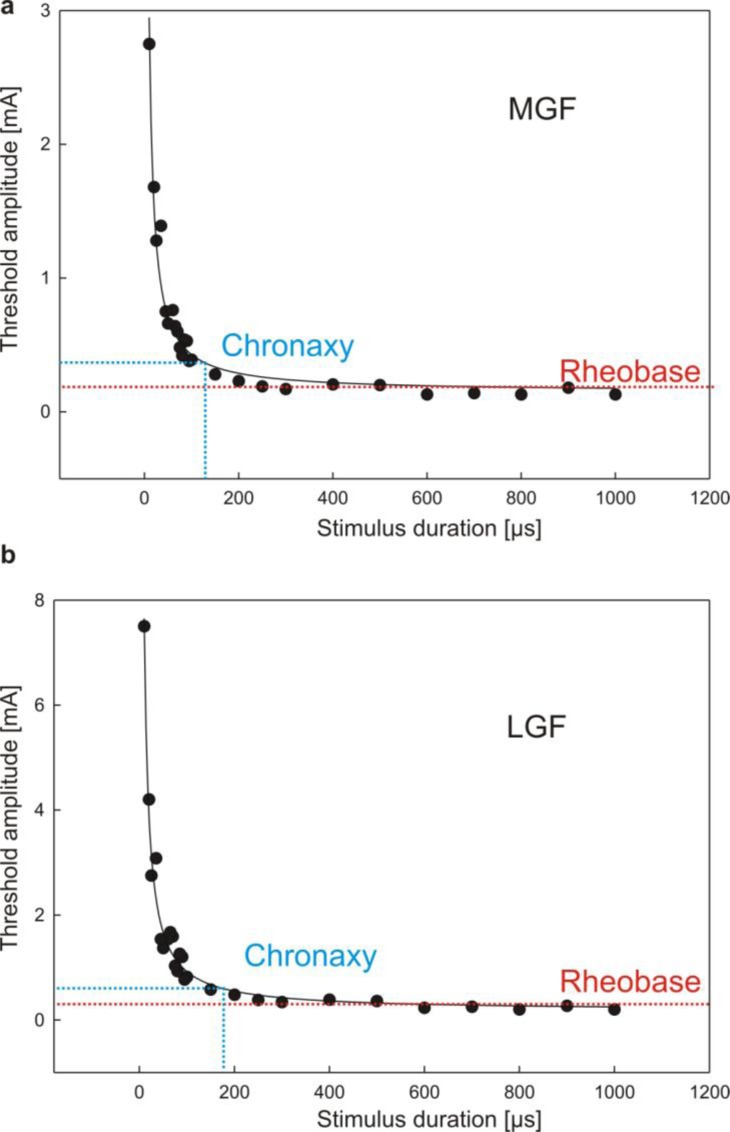
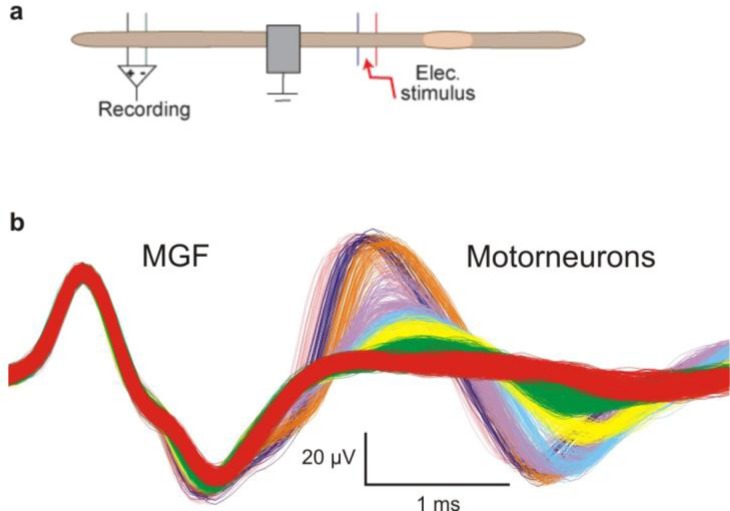
References
-
- Bear MF, Connors BW, Paradiso MA. Neuroscience, exploring the brain. 3rd ed. Baltimore, MD: Lippincott Williams and Wilkins; 2007.
-
- Bullock TH. Functional organization of the giant fibre system of Lumbricus. J Neurophysiol. 1945;8:54–71.
-
- Bullock TH, Horridge GA. Structure and function in the nervous systems of invertebrates. San Francisco, CA: Freeman and Company; 1965.
LinkOut - more resources
Full Text Sources
Research Materials