

Viruses and prions of Saccharomyces cerevisiae
- PMID: 23498901
- PMCID: PMC4141569
- DOI: 10.1016/B978-0-12-394315-6.00001-5
Viruses and prions of Saccharomyces cerevisiae
Abstract
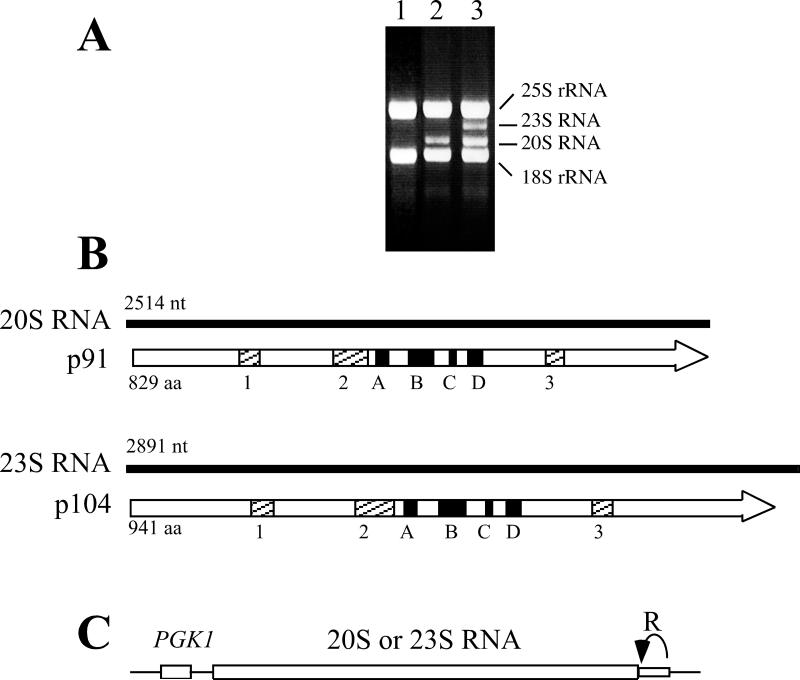
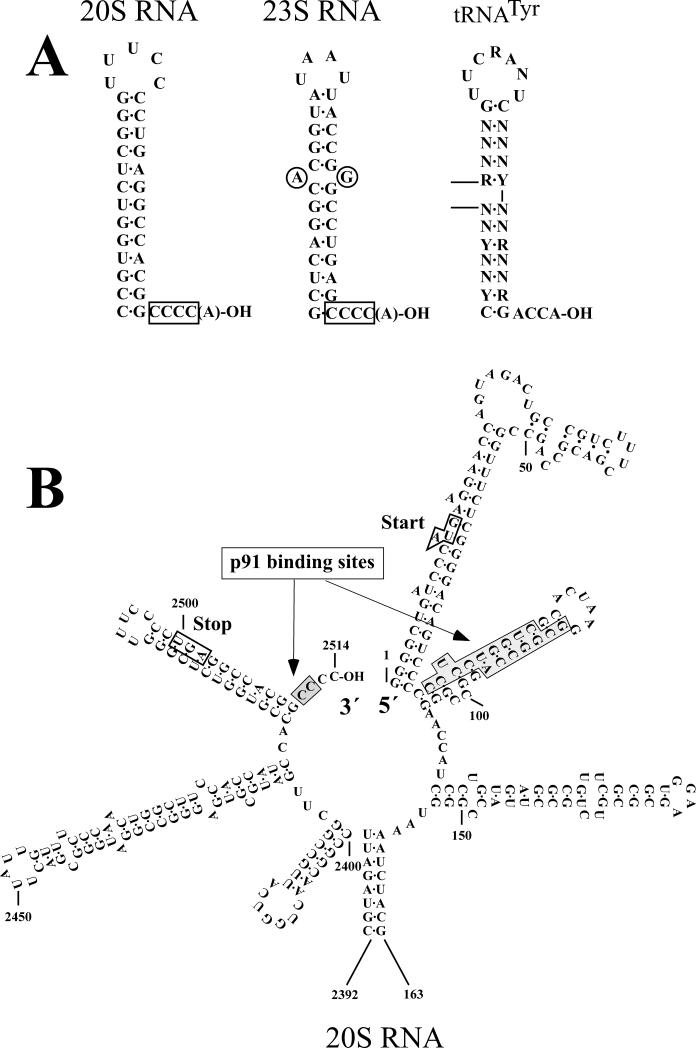
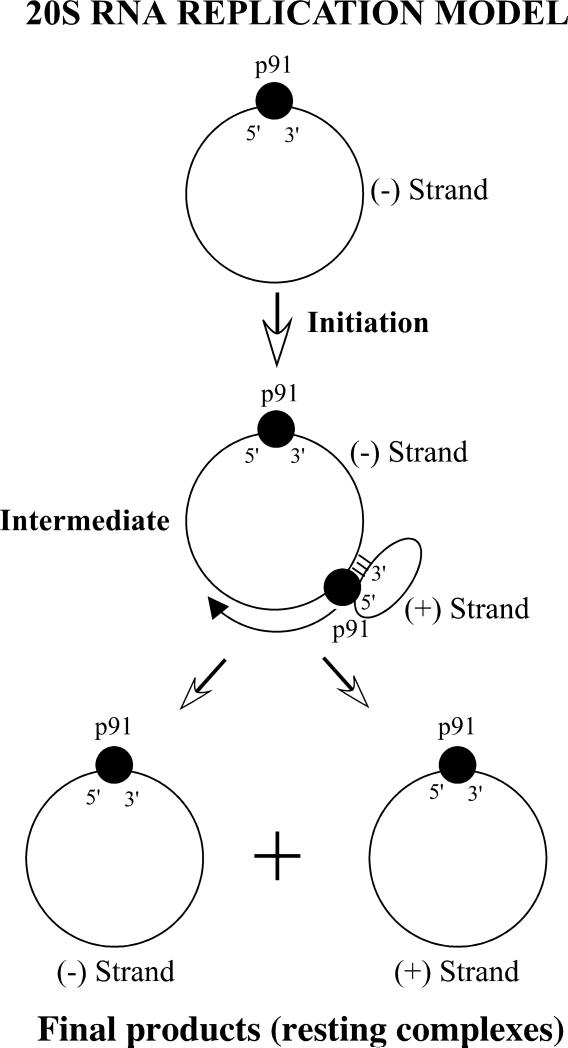
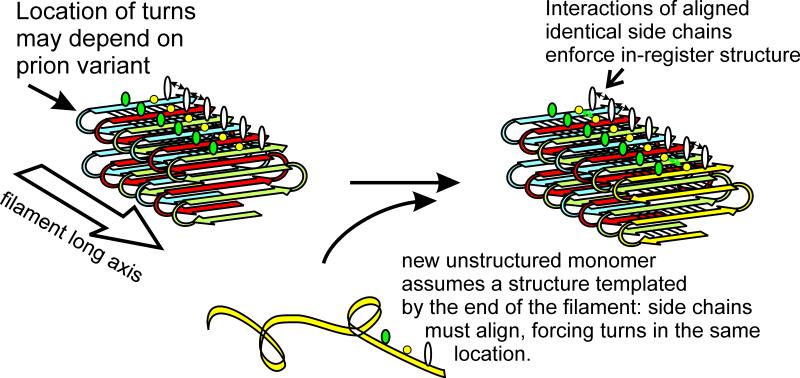
Saccharomyces cerevisiae has been a key experimental organism for the study of infectious diseases, including dsRNA viruses, ssRNA viruses, and prions. Studies of the mechanisms of virus and prion replication, virus structure, and structure of the amyloid filaments that are the basis of yeast prions have been at the forefront of such studies in these classes of infectious entities. Yeast has been particularly useful in defining the interactions of the infectious elements with cellular components: chromosomally encoded proteins necessary for blocking the propagation of the viruses and prions, and proteins involved in the expression of viral components. Here, we emphasize the L-A dsRNA virus and its killer-toxin-encoding satellites, the 20S and 23S ssRNA naked viruses, and the several infectious proteins (prions) of yeast.
Copyright © 2013 Elsevier Inc. All rights reserved.
Figures
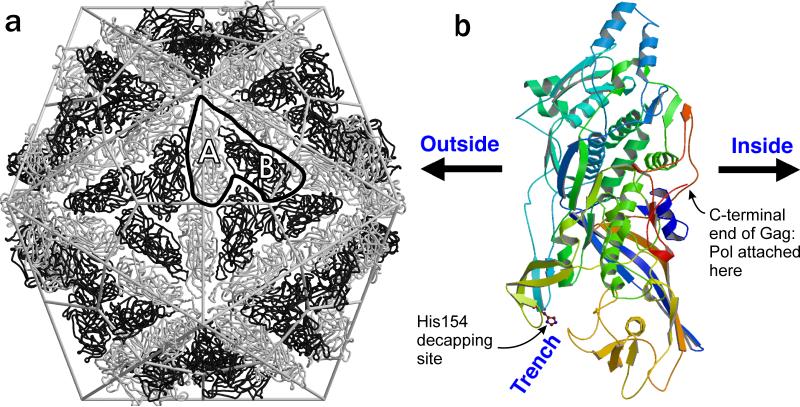
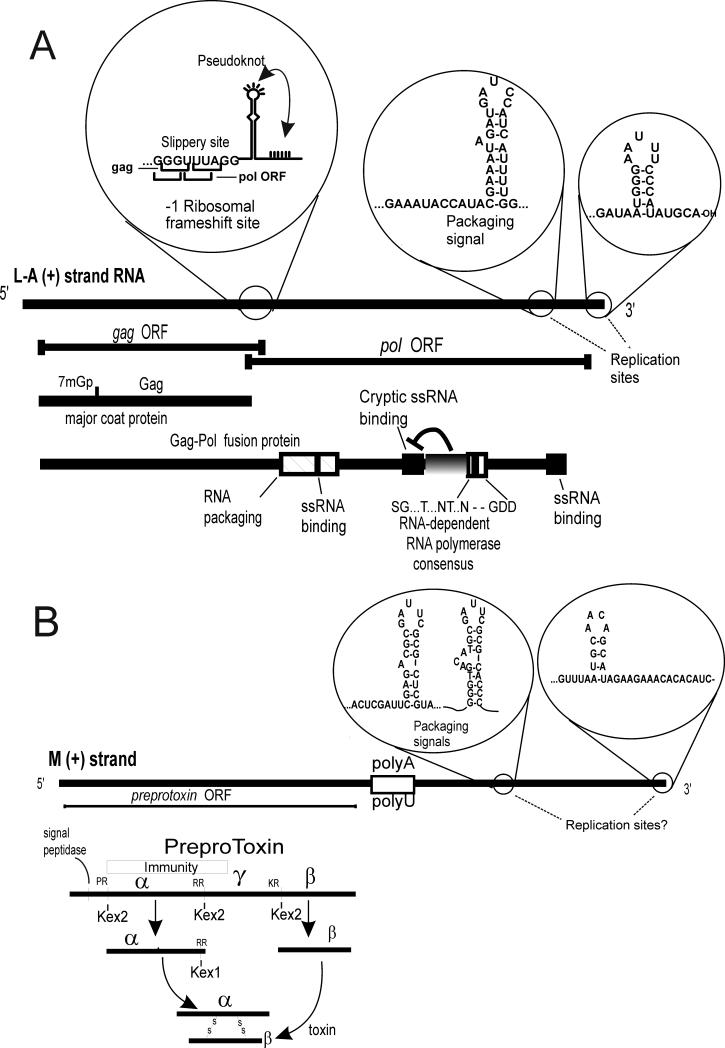
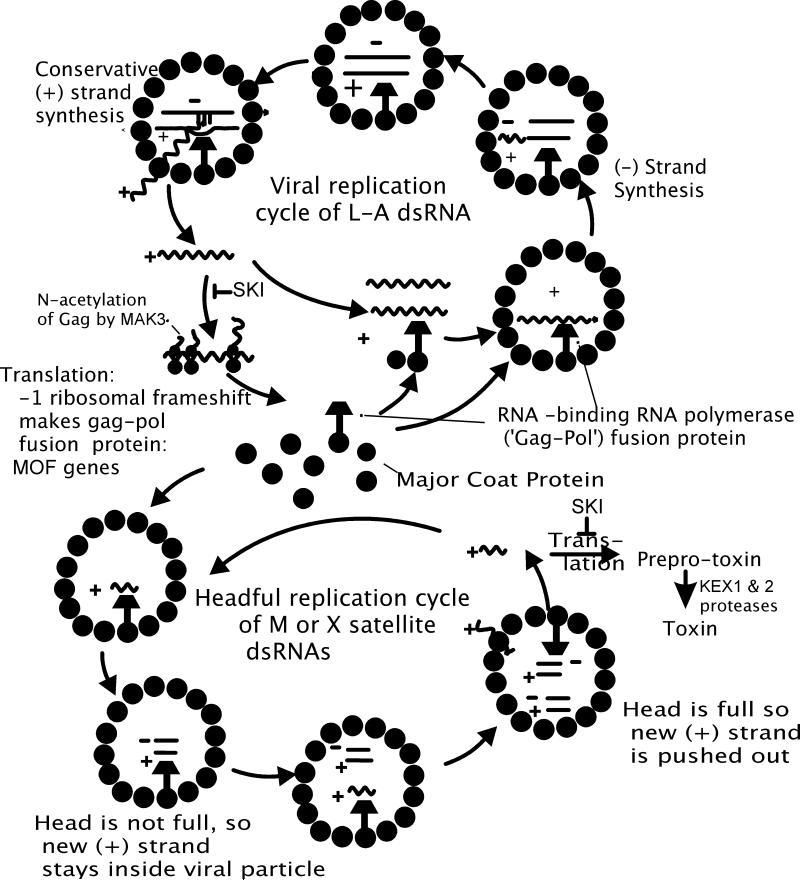
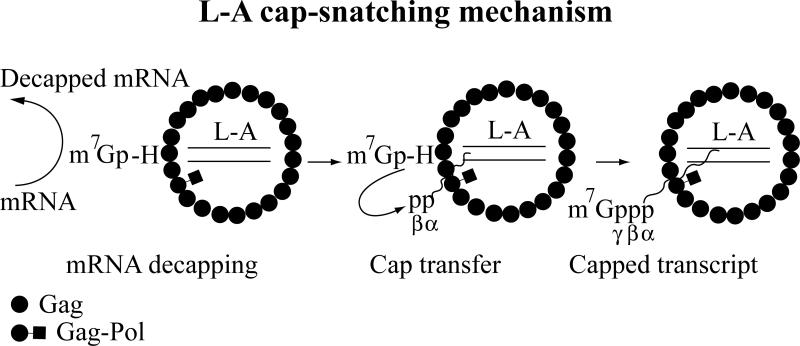
References
-
- Bai M, Zhou JM, Perrett S. The yeast prion protein Ure2 shows glutathione peroxidase activity in both native and fibrillar forms. J Biol Chem. 2004;279:50025–30. - PubMed
-
- Baxa U, Cheng N, Winkler DC, Chiu TK, Davies DR, Sharma D, Inouye H, Kirschner DA, Wickner RB, Steven AC. Filaments of the Ure2p prion protein have a cross-beta core structure. J Struct Biol. 2005;150:170–9. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
