

Review
doi: 10.1016/j.cell.2013.02.034.
Dynamic integration of splicing within gene regulatory pathways
Affiliations
- PMID: 23498935
- PMCID: PMC3642998
- DOI: 10.1016/j.cell.2013.02.034
Item in Clipboard
Review
Dynamic integration of splicing within gene regulatory pathways
Cell.
.
Abstract
Precursor mRNA splicing is one of the most highly regulated processes in metazoan species. In addition to generating vast repertoires of RNAs and proteins, splicing has a profound impact on other gene regulatory layers, including mRNA transcription, turnover, transport, and translation. Conversely, factors regulating chromatin and transcription complexes impact the splicing process. This extensive crosstalk between gene regulatory layers takes advantage of dynamic spatial, physical, and temporal organizational properties of the cell nucleus, and further emphasizes the importance of developing a multidimensional understanding of splicing control.
Copyright © 2013 Elsevier Inc. All rights reserved.
Figures
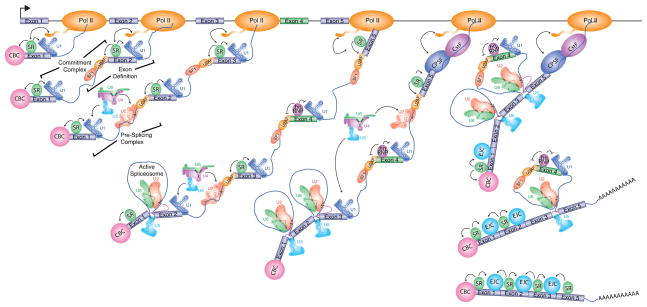
Co-transcriptional spliceosome assembly initiates with the binding of U1 snRNP to the 5′ splice site, which is enhanced by exon-bound SR proteins and, for the first exon, the Cap Binding Complex (CBC). A cross-intron commitment complex is formed upon association of U2 snRNP auxiliary factor (U2AF) with the 3′ splice site and adjacent intronic polypyrimidine tract, and branch point binding protein (BBP/SF1) with the branch site. Bridging interactions between these factors across internal exons, or “exon-definition”, occurs within the commitment complex. Transition from a commitment complex to a pre-splicing complex entails communication between 5′ and 3′ splice sites, and the addition of U2 snRNP to the branch site along with numerous additional proteins (not shown). Subsequent association of U4/U6/U5 tri-snRNP, together with still more protein factors, and dynamic remodeling of RNA-protein, protein-protein, and RNA-RNA interactions, ultimately leads to formation of the catalytically-active spliceosome. The two trans-esterification steps of splicing yield the excised intron in the form of the characteristic branched “lariat” structure, and the ligated exons that form mature mRNA. The assembly of most splicing factors and splicing of constitutive introns is thought to occur co-transcriptionally, while splicing of regulated alternative introns often occurs post-transcriptionally. In the example shown, exon 4 is a regulated alternative exon controlled by an hnRNP protein, which prevents the splicing factors bound to flanking splice sites from engaging in productive interactions and therefore promotes exon skipping. At terminal exons (Exon 5), interactions between the splicing factors bound to the upstream 3' splice site and the exon interact with components of the cleavage and polyadenlyation machinery (CPSF and CstF are shown; see also Figure 4A). The association of the splicing factors with the pre-mRNA is enhanced throughout the transcription process by interactions with the C-terminal domain of RNA polymerase II. The Exon-Junction complex (EJC) is recruited upstream of splice junctions upon splicing. The EJC and SR proteins mutually stabilize one another to generate the mature mRNP, which is then exported to the cytoplasm.
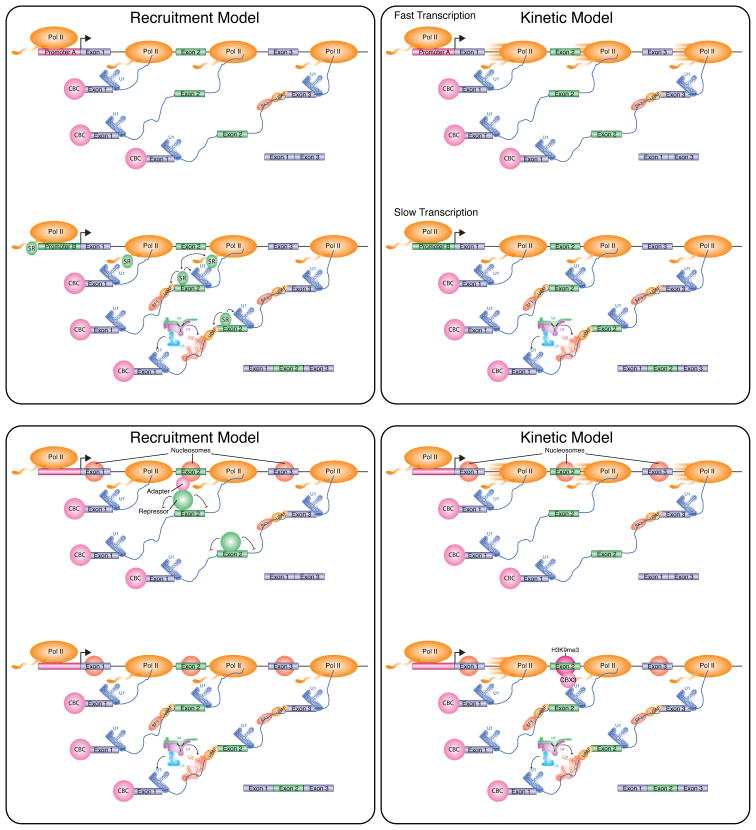
(Top Left) Promoter recruitment model. Different promoters differentially recruit splicing factors to the transcription complex. At promoters which fail to recruit a key splicing factor (shown as an SR protein), the regulated alternative exon (Exon 2) will be skipped, while genes containing promoters that recruit the splicing factor will include exon 2. (Top Right) Promoter-directed kinetic model. Different promoters assemble transcription complexes capable of different transcription elongation rates. At promoters that assemble fast transcription elongation complexes, the regulated alternative exon (Exon 2) will be skipped, while genes containing promoters that assemble slow elongation complexes will include exon 2. This model requires that the alternative exon contains weak 3′ and/or 5′ splice sites in order to be skipped when the gene is rapidly transcribed. (Bottom Left) Chromatin-mediated recruitment model. The splicing of an alternative exon can be regulated by the chromatin-mediated recruitment of a splicing repressor. In cells that skip the exon, an adapter protein associates with the nucleosome assembled at the alternative exon, which in turn recruits a splicing repressor. In cells that include the alternative exon, the adaptor protein and/or repressor are not expressed, or the nucleosome at the regulated alternative exon is not modified and therefore cannot recruit the repressor. Similar to this model, a nucleosome-associated adaptor protein may also function to recruit a splicing activator, as proposed for Psip1/Ledgf (Pradeepa et al., 2012) (Bottom Right) Chromatin-mediated kinetic model. The splicing of an alternative exon can be regulated by a chromatin-mediated change in the rate of transcription elongation. Unmodified nucleosomes can be transcribed rapidly, resulting in skipping of the regulated alternative exon. In cells where the nucleosome assembled on exon 2 has an H3K9me3 mark, CBX3 interacts with the modified nucleosome, slows down the transcription elongation complex, and enhances splicing of the regulated alternative exon.
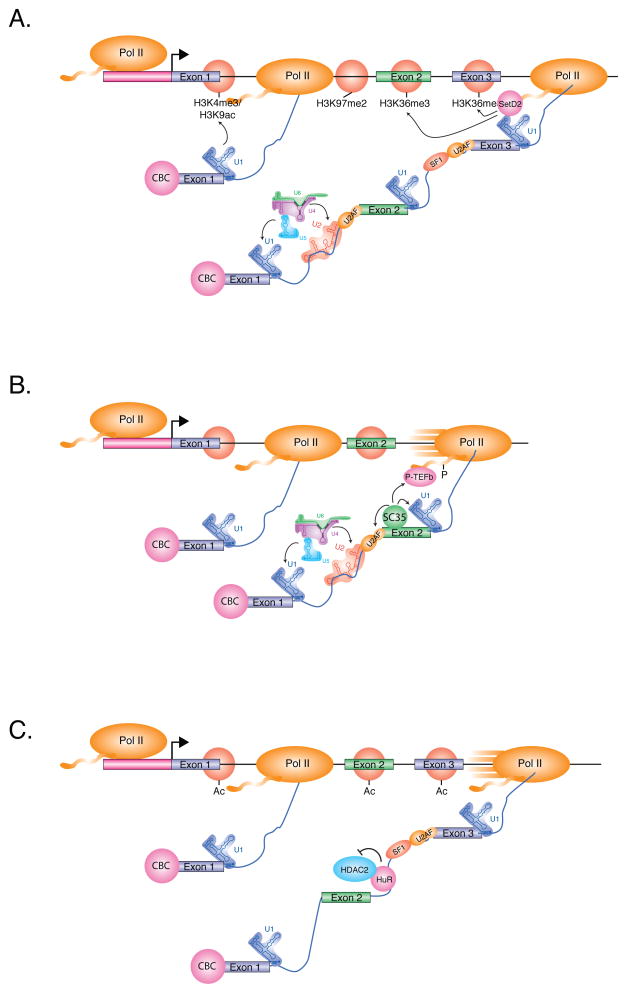
(A) Splicing enhances transcription-associated histone modification. Splicing of the first intron enhances transcription initiation and stabilizes promoter-associated marks, including H3K4me3 and H3K9ac, near the 5' splice site of exon 1. Splicing may also facilitate a transition between the elongation-associated marks H3K79me2 and H3K36me3 at the 3' splice site of the first intron. Internal exons are particularly enriched for H3K36me3-modified nucleosomes, due in part to splicing-increased nucleosome occupancy and action of the histone methyltransferase SETD2 associated with elongating Pol II. These marks may also serve to reinforce splicing patterns of nascent pre-mRNA. (B) The SR protein SRSF2/SC35, which regulates splicing of alternative exons, also enhances transcription elongation by recruiting P-TEFb. P-TEFb phosphorylates the Pol II CTD at Serine 2, which enhances the rate of transcription elongation. (C) The Hu family of splicing regulators bind to AU-rich sequences within introns and repress the splicing of regulated alternative exons. Shown here, HuR interacts with and represses the activity of the histone deacetylase, HDAC2, which stabilizes nearby acetylated nucleosomes. Acetylated nucleosomes may enhance the rate of transcription elongation, and consequently, promote the skipping of exons with weak splice sites.
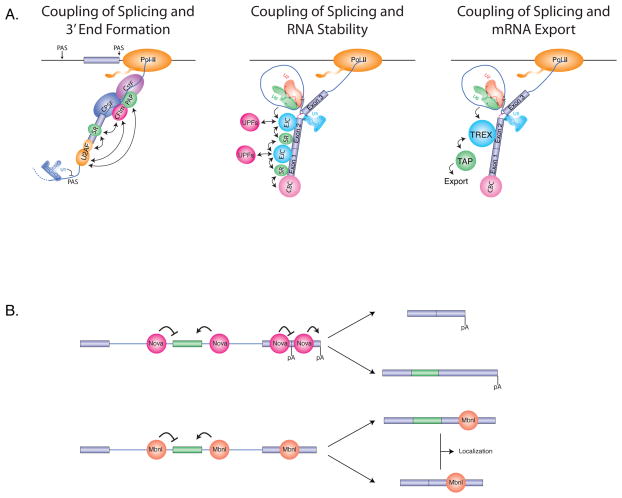
(A) Coupling connections between splicing and 3′ end formation, RNA stability, and mRNA export. Splicing and 3'-end formation are coupled by interactions between exon-bound SR proteins and the cleavage and polyadenylation factor CFIm, and between U2AF and both CFIm and PAP. Cryptic upstream polyadenlyation sites (PAS) are suppressed by U1 snRNP (left). Splicing is impacts RNA stability by interactions between SR proteins and the EJC, which in turn interacts with the UPF proteins involved in nonsense mediated decay (NMD) (middle). Splicing influences mRNA export through the splicing-dependent recruitment of the TREX complex, which in turn interacts with the RNA export factor TAP. (B). Multi-tasking roles of RBPs in splicing and alternative polyadenylation, RNA export and RNA transport. (Top) The Nova RNA binding proteins have been shown to not only regulate alternative splicing, but also alternative polyadenylation (pA). Both of these processes are modulated in a position-dependent manner with some binding locations promoting splicing and polyadenylation, and other locations repressing these processes. The result of this regulation is the generation of mRNAs with different exons and 3' UTR sequences (Bottom) Similarly, Mbnl RNA binding proteins impact alternative splicing in a position-dependent manner, and to bind to 3' UTRs, where they function to control subcellular mRNA localization.
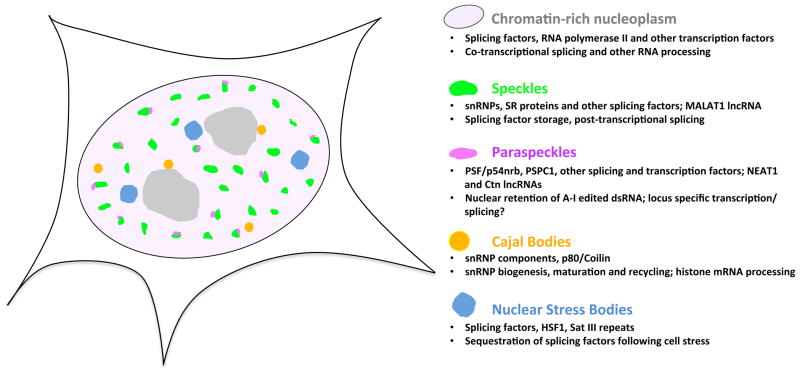
Major nuclear domains enriched in splicing and other factors in the mammalian cell nucleus are depicted, with known and putative roles indicated. Gray areas indicate nucleoli.
References
-
- Ameur A, Zaghlool A, Halvardson J, Wetterbom A, Gyllensten U, Cavelier L, Feuk L. Total RNA sequencing reveals nascent transcription and widespread co-transcriptional splicing in the human brain. Nat Struct Mol Biol. 2011;18:1435–1440. - PubMed
-
- Ameyar-Zazoua M, Rachez C, Souidi M, Robin P, Fritsch L, Young R, Morozova N, Fenouil R, Descostes N, Andrau JC, et al. Argonaute proteins couple chromatin silencing to alternative splicing. Nat Struct Mol Biol. 2012;19:998–1004. - PubMed
-
- Amit M, Donyo M, Hollander D, Goren A, Kim E, Gelfman S, Lev-Maor G, Burstein D, Schwartz S, Postolsky B, et al. Differential GC content between exons and introns establishes distinct strategies of splice-site recognition. Cell Rep. 2012;1:543–556. - PubMed
-
- Auboeuf D, Honig A, Berget SM, O'Malley BW. Coordinate regulation of transcription and splicing by steroid receptor coregulators. Science. 2002;298:416–419. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
