

Epigenetics of reprogramming to induced pluripotency
- PMID: 23498940
- PMCID: PMC3602907
- DOI: 10.1016/j.cell.2013.02.043
Epigenetics of reprogramming to induced pluripotency
Abstract
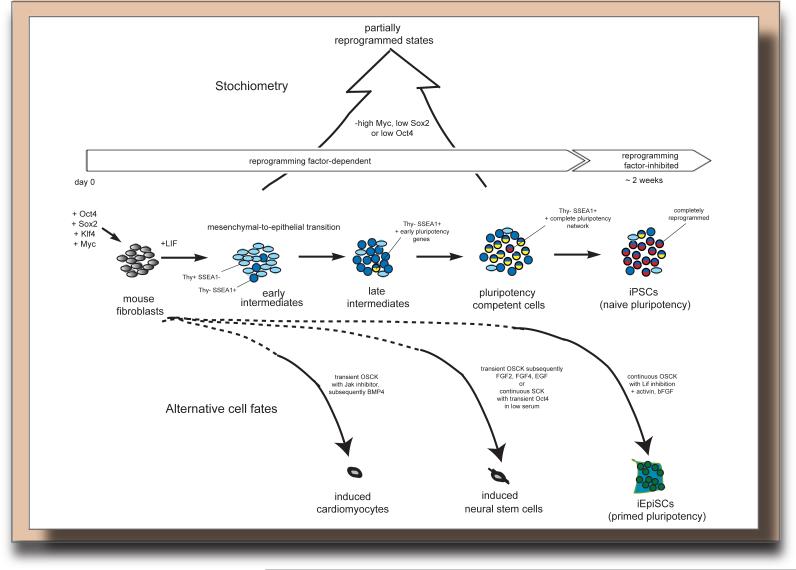
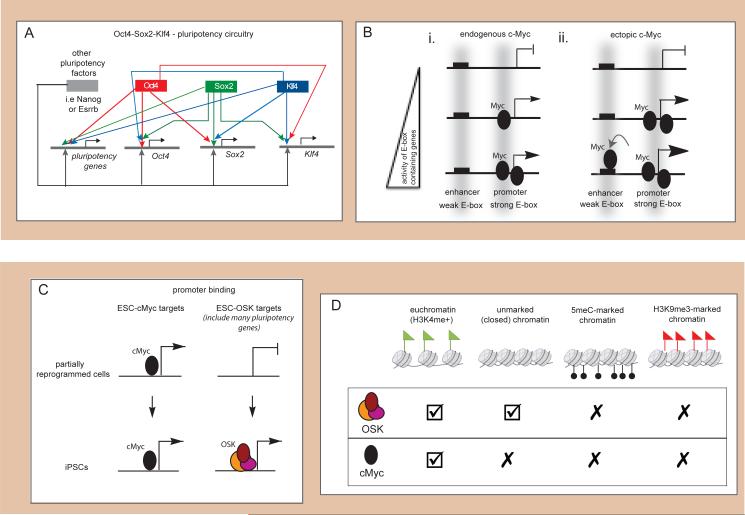
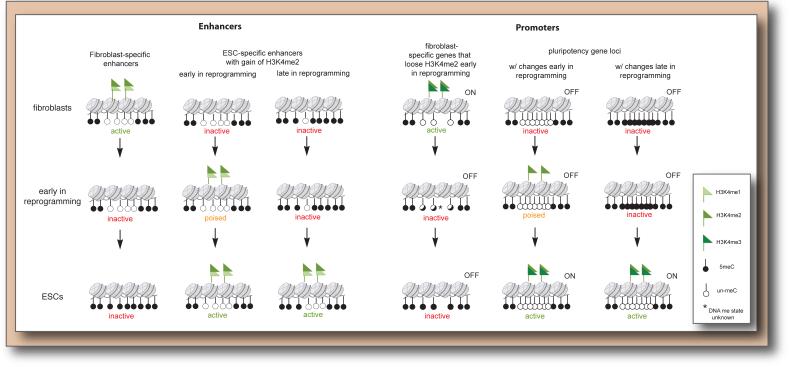
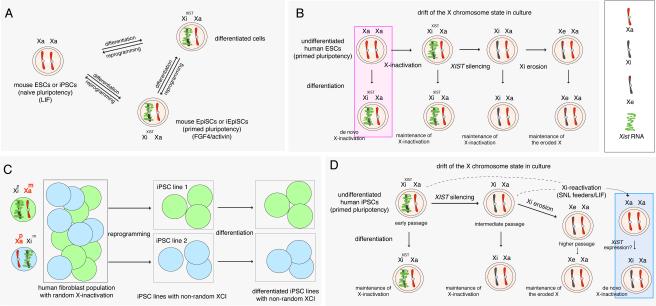
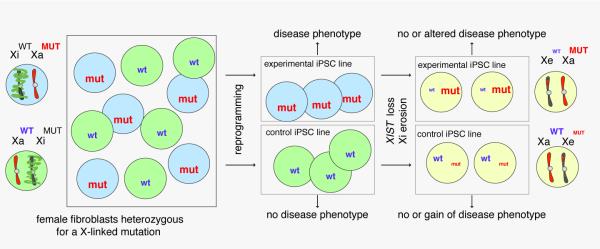
Reprogramming to induced pluripotent stem cells (iPSCs) proceeds in a stepwise manner with reprogramming factor binding, transcription, and chromatin states changing during transitions. Evidence is emerging that epigenetic priming events early in the process may be critical for pluripotency induction later. Chromatin and its regulators are important controllers of reprogramming, and reprogramming factor levels, stoichiometry, and extracellular conditions influence the outcome. The rapid progress in characterizing reprogramming is benefiting applications of iPSCs and is already enabling the rational design of novel reprogramming factor cocktails. However, recent studies have also uncovered an epigenetic instability of the X chromosome in human iPSCs that warrants careful consideration.
Copyright © 2013 Elsevier Inc. All rights reserved.
Figures
References
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
