

Incoming RNA virus nucleocapsids containing a 5'-triphosphorylated genome activate RIG-I and antiviral signaling
- PMID: 23498958
- PMCID: PMC5515363
- DOI: 10.1016/j.chom.2013.01.012
Incoming RNA virus nucleocapsids containing a 5'-triphosphorylated genome activate RIG-I and antiviral signaling
Abstract
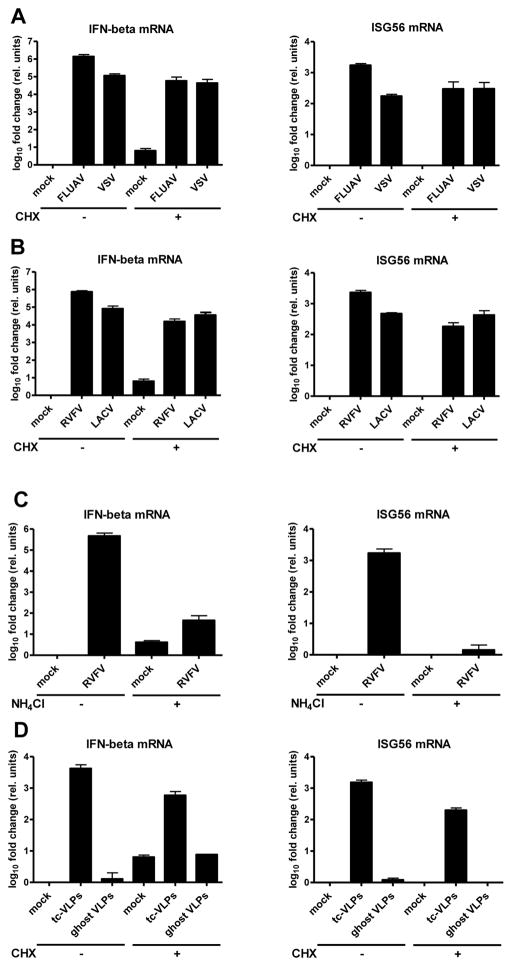
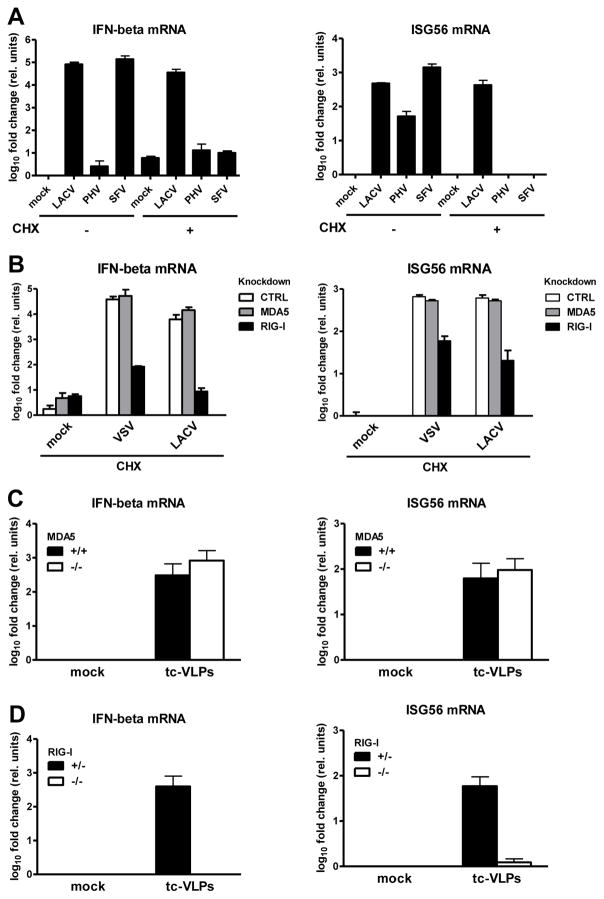
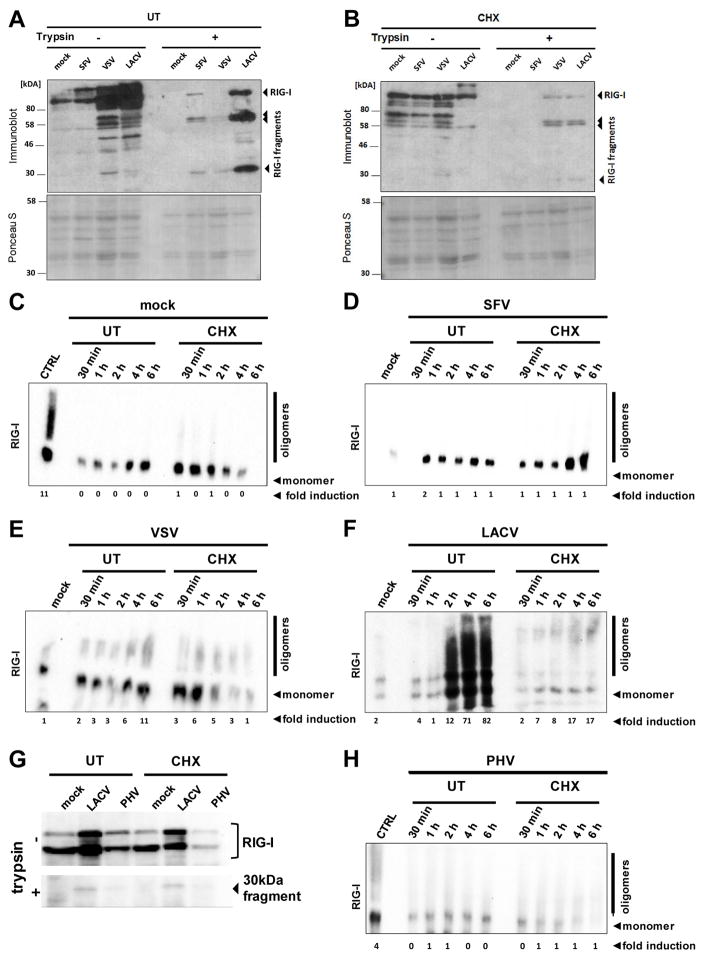
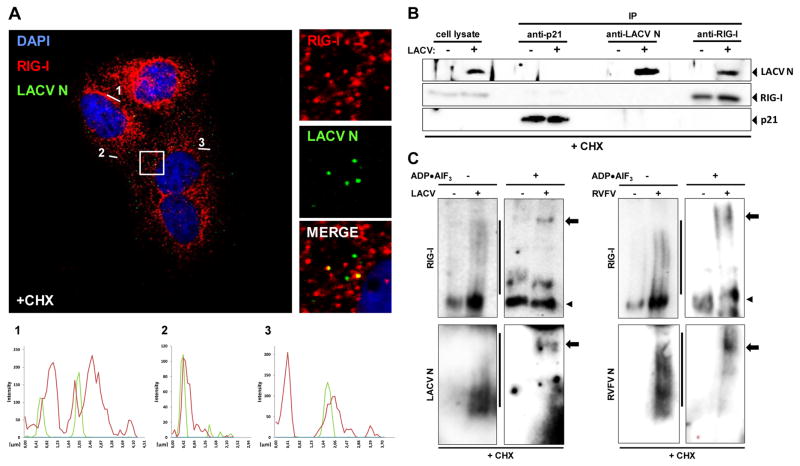
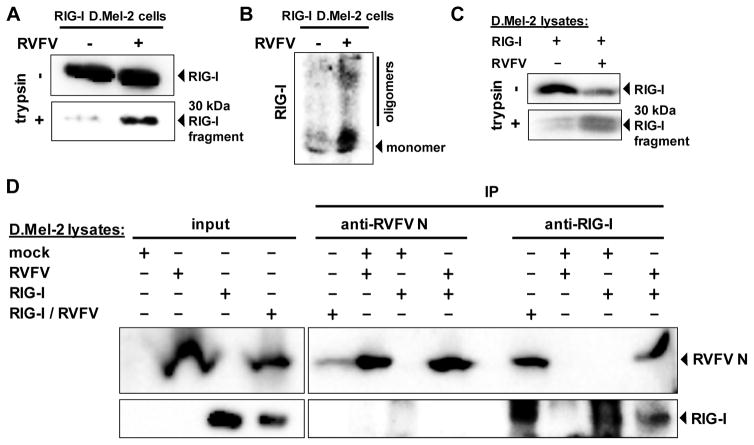
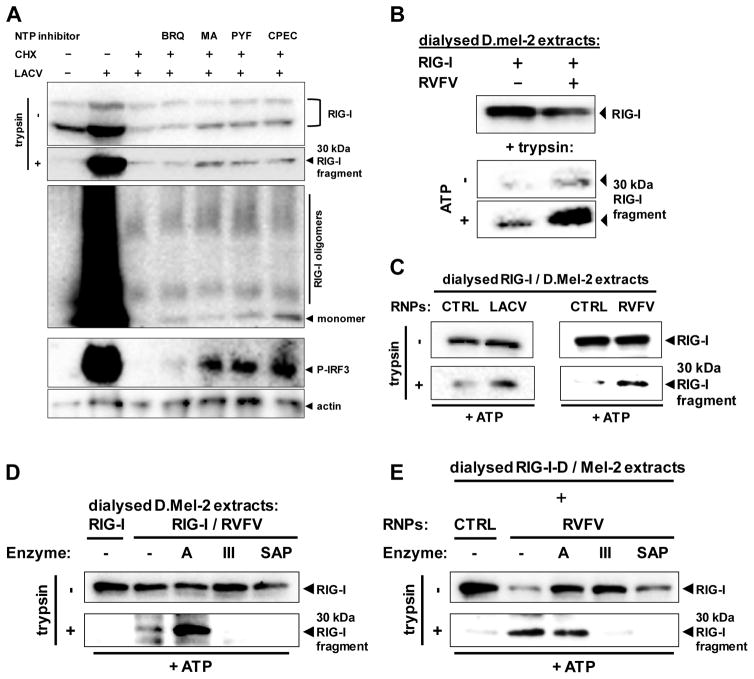
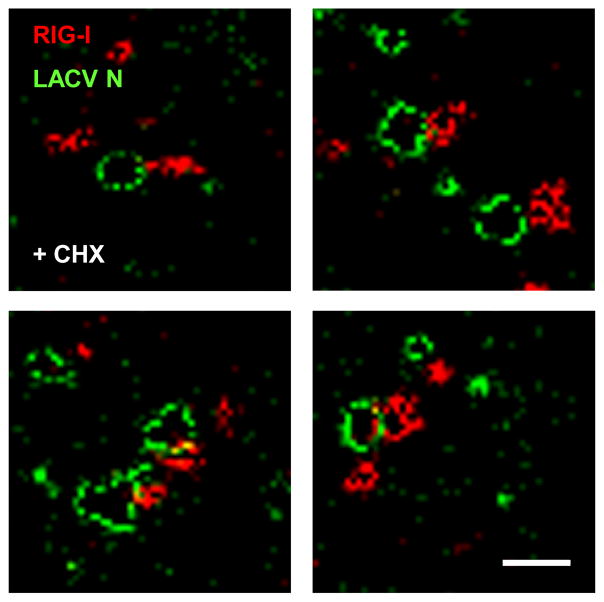
Host defense to RNA viruses depends on rapid intracellular recognition of viral RNA by two cytoplasmic RNA helicases: RIG-I and MDA5. RNA transfection experiments indicate that RIG-I responds to naked double-stranded RNAs (dsRNAs) with a triphosphorylated 5' (5'ppp) terminus. However, the identity of the RIG-I stimulating viral structures in an authentic infection context remains unresolved. We show that incoming viral nucleocapsids containing a 5'ppp dsRNA "panhandle" structure trigger antiviral signaling that commences with RIG-I, is mediated through the adaptor protein MAVS, and terminates with transcription factor IRF-3. Independent of mammalian cofactors or viral polymerase activity, RIG-I bound to viral nucleocapsids, underwent a conformational switch, and homo-oligomerized. Enzymatic probing and superresolution microscopy suggest that RIG-I interacts with the panhandle structure of the viral nucleocapsids. These results define cytoplasmic entry of nucleocapsids as the proximal RIG-I-sensitive step during infection and establish viral nucleocapsids with a 5'ppp dsRNA panhandle as a RIG-I activator.
Copyright © 2013 Elsevier Inc. All rights reserved.
Figures
Comment in
-
RIG-I goes beyond naked recognition.Cell Host Microbe. 2013 Mar 13;13(3):247-9. doi: 10.1016/j.chom.2013.02.012. Cell Host Microbe. 2013. PMID: 23498950 Free PMC article.
References
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Miscellaneous
