

Neurodegeneration-associated protein fragments as short-lived substrates of the N-end rule pathway
- PMID: 23499006
- PMCID: PMC3640747
- DOI: 10.1016/j.molcel.2013.02.009
Neurodegeneration-associated protein fragments as short-lived substrates of the N-end rule pathway
Abstract
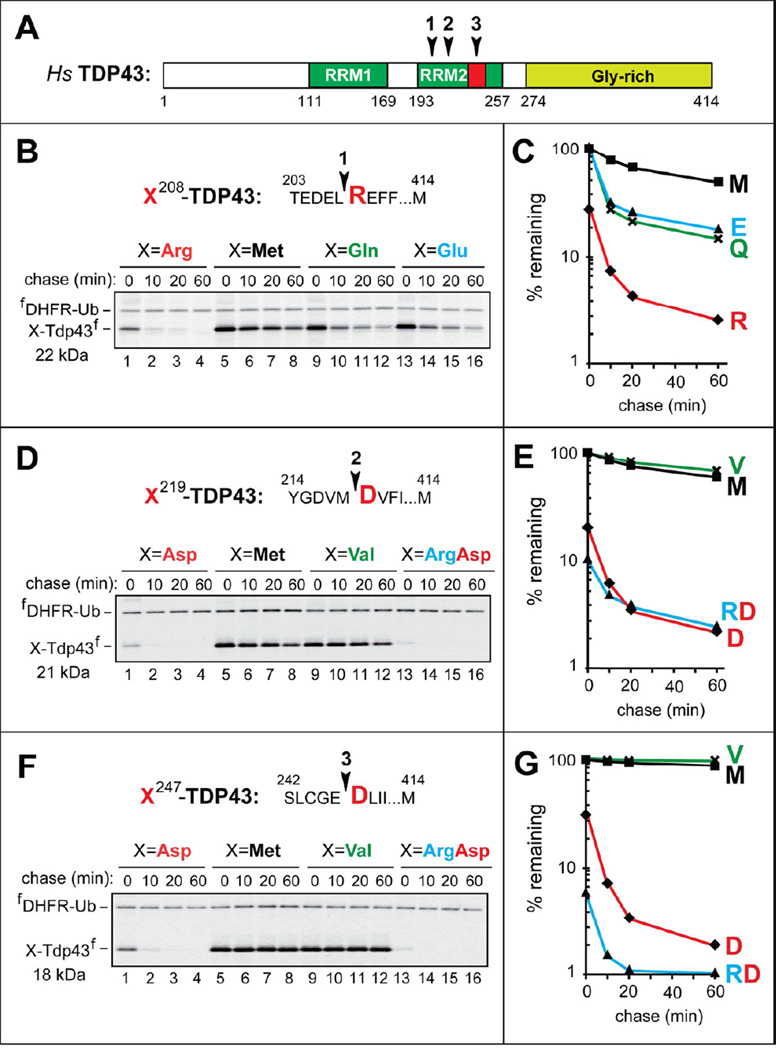
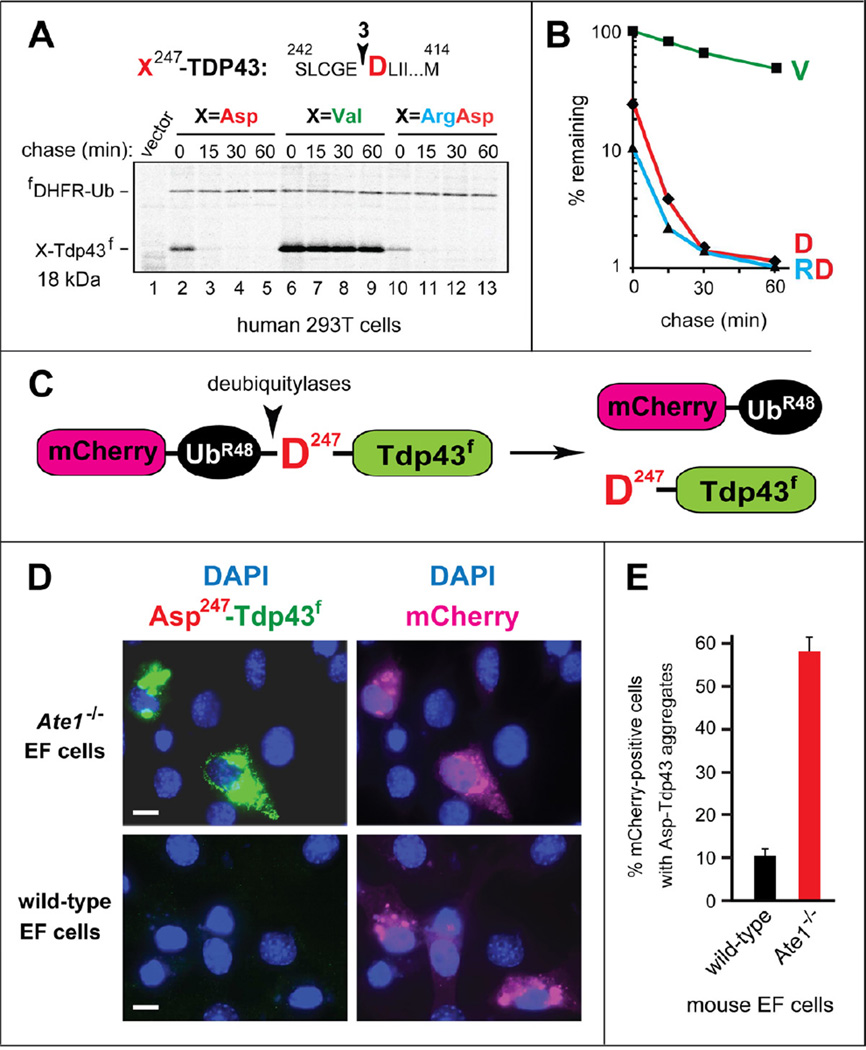
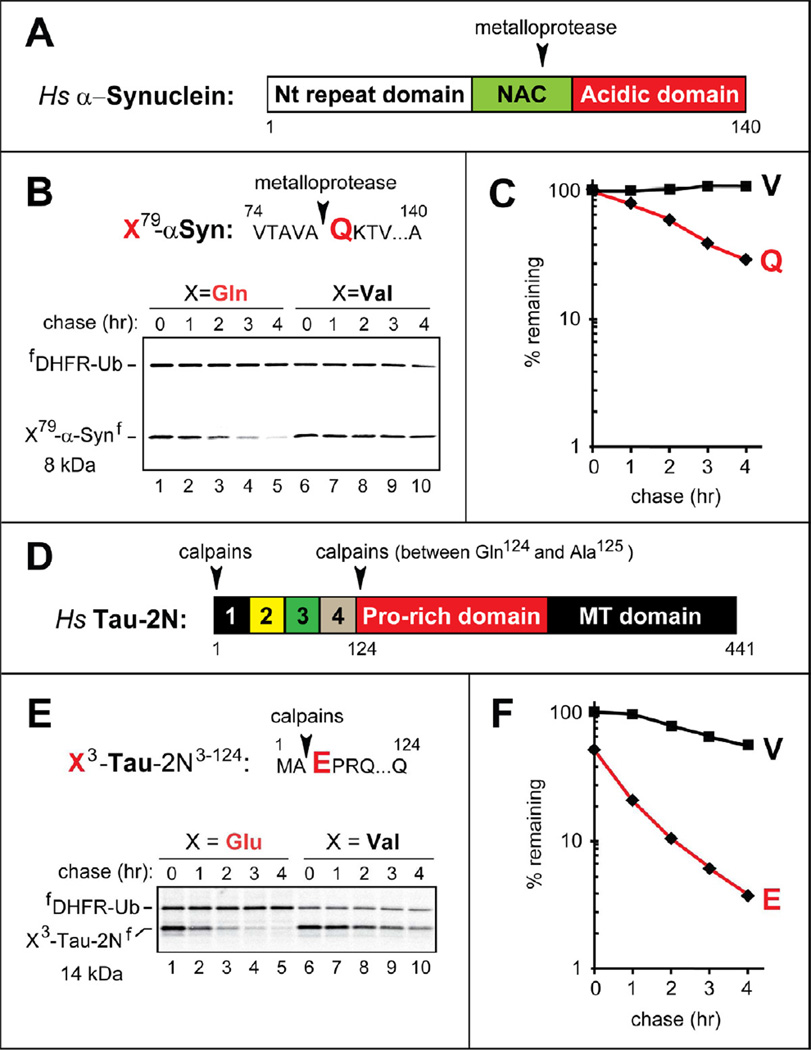
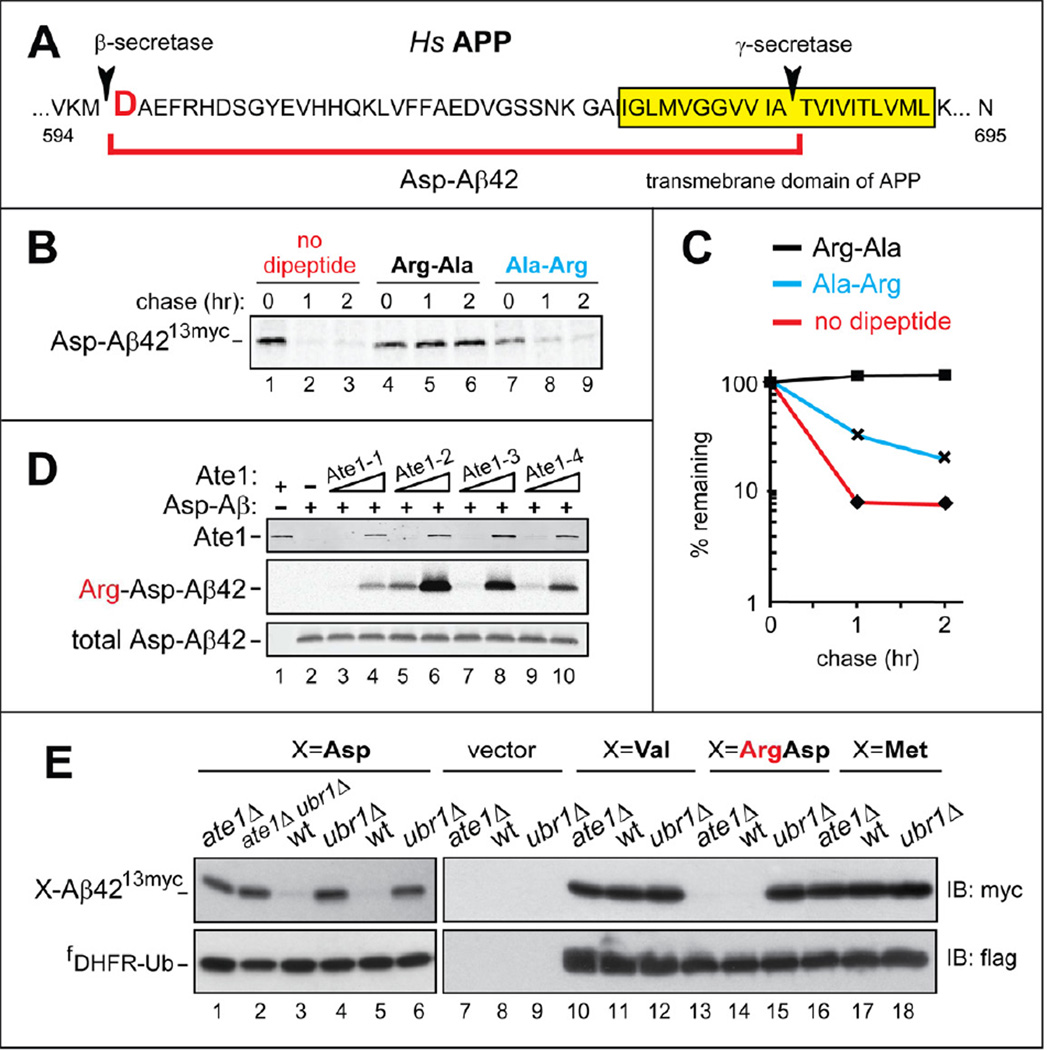
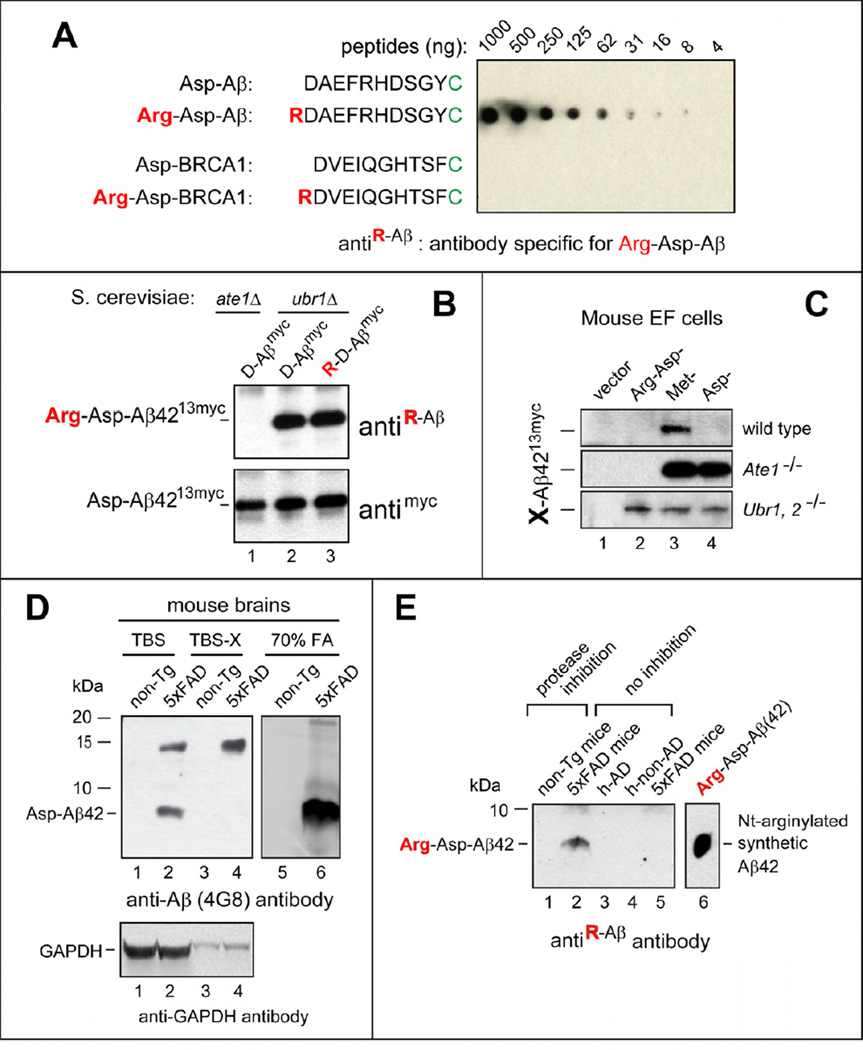
Protein aggregates are a common feature of neurodegenerative syndromes. Specific protein fragments were found to be aggregated in disorders including Alzheimer's disease, amyotrophic lateral sclerosis, and Parkinson's disease. Here, we show that the natural C-terminal fragments of Tau, TDP43, and α-synuclein are short-lived substrates of the Arg/N-end rule pathway, a processive proteolytic system that targets proteins bearing "destabilizing" N-terminal residues. Furthermore, a natural TDP43 fragment is shown to be metabolically stabilized in Ate1(-/-) fibroblasts that lack the arginylation branch of the Arg/N-end rule pathway, leading to accumulation and aggregation of this fragment. We also found that a fraction of Aβ42, the Alzheimer's disease-associated fragment of APP, is N-terminally arginylated in the brains of 5xFAD mice and is degraded by the Arg/N-end rule pathway. The discovery that neurodegeneration-associated natural fragments of TDP43, Tau, α-synuclein, and APP can be selectively destroyed by the Arg/N-end rule pathway suggests that this pathway counteracts neurodegeneration.
Copyright © 2013 Elsevier Inc. All rights reserved.
Figures
References
-
- Balch WE, Morimoto RI, Dillin A, Kelly JW. Adapting proteostasis for disease intervention. Science. 2008;319:916–919. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
Research Materials
Miscellaneous
