

Cohesin-dependent association of scc2/4 with the centromere initiates pericentromeric cohesion establishment
- PMID: 23499533
- PMCID: PMC3627958
- DOI: 10.1016/j.cub.2013.02.022
Cohesin-dependent association of scc2/4 with the centromere initiates pericentromeric cohesion establishment
Abstract
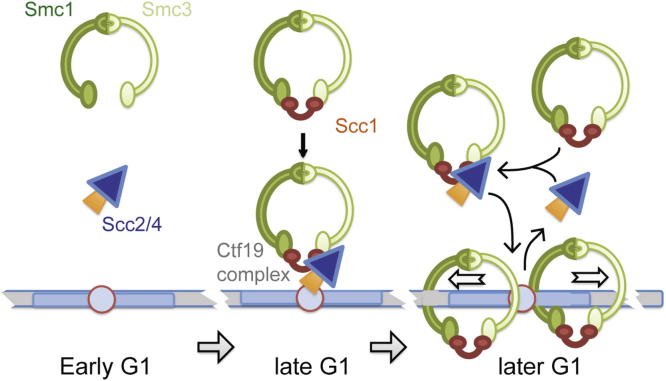
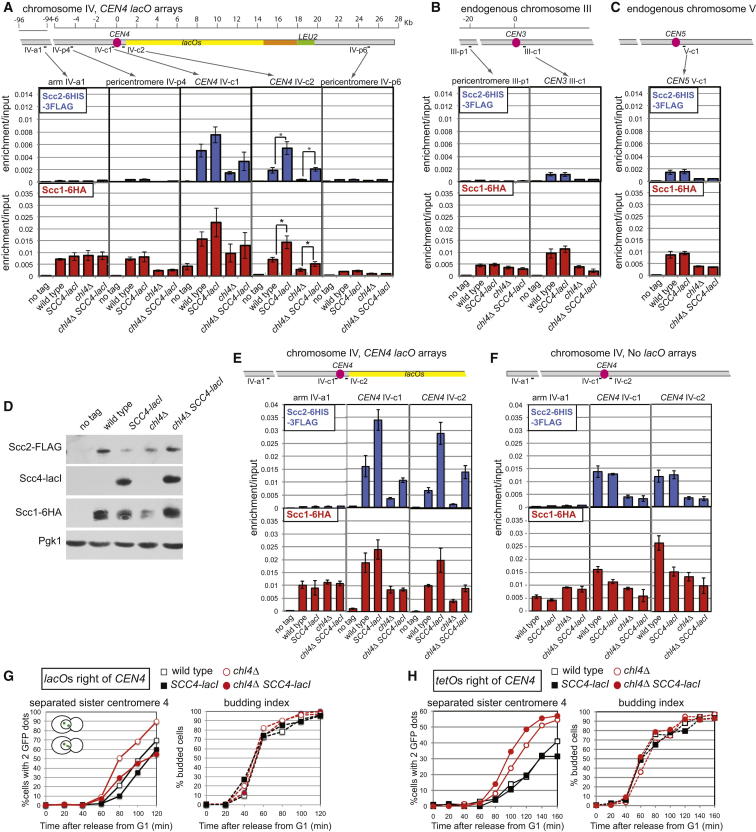
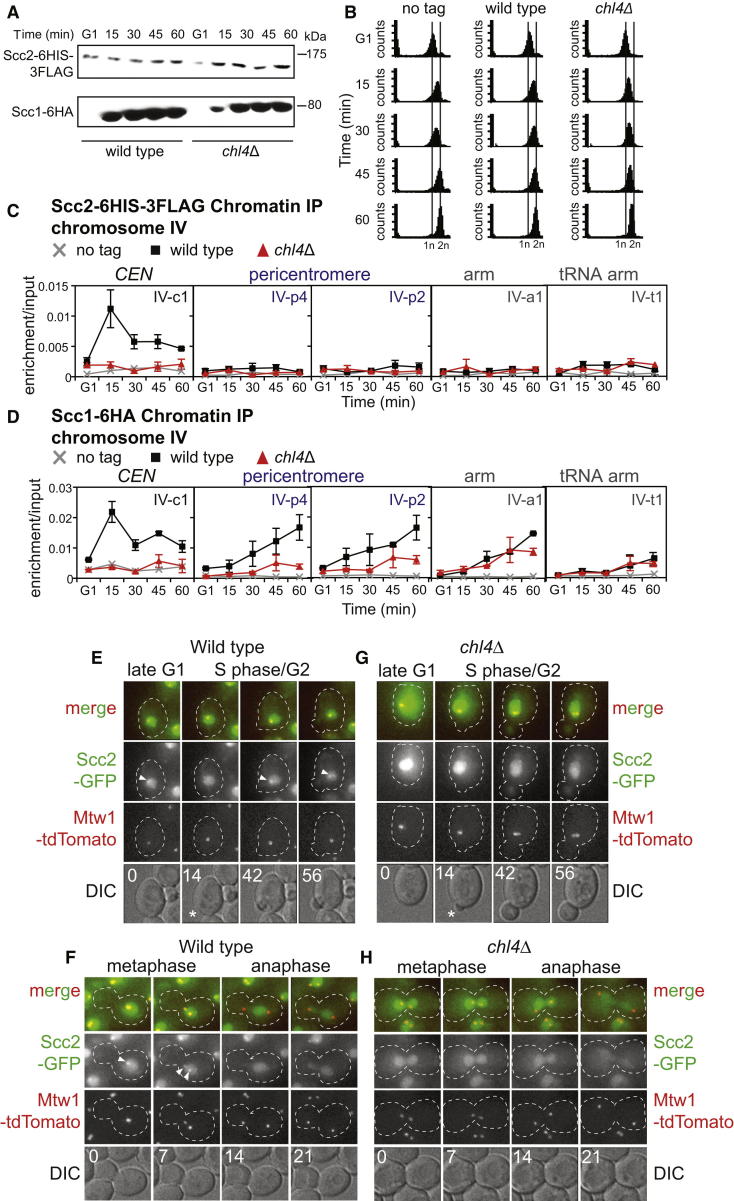
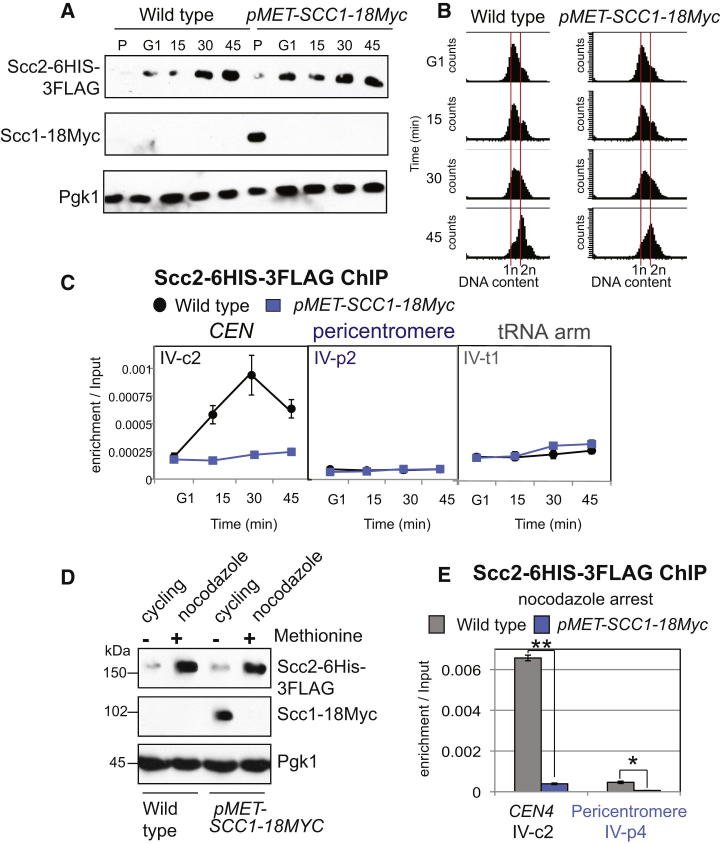
Cohesin is a conserved ring-shaped multiprotein complex that participates in chromosome segregation, DNA repair, and transcriptional regulation [1, 2]. Cohesin loading onto chromosomes universally requires the Scc2/4 "loader" complex (also called NippedBL/Mau2), mutations in which cause the developmental disorder Cornelia de Lange syndrome in humans [1-9]. Cohesin is most concentrated in the pericentromere, the region surrounding the centromere [10-15]. Enriched pericentromeric cohesin requires the Ctf19 kinetochore subcomplex in budding yeast [16-18]. Here, we uncover the spatial and temporal determinants for Scc2/4 centromere association. We demonstrate that the critical role of the Ctf19 complex is to enable Scc2/4 association with centromeres, through which cohesin loads and spreads onto the adjacent pericentromere. We show that, unexpectedly, Scc2 association with centromeres depends on cohesin itself. The absence of the Scc1/Mcd1/Rad21 cohesin subunit precludes Scc2 association with centromeres from anaphase until late G1. Expression of SCC1 is both necessary and sufficient for the binding of cohesin to its loader, the association of Scc2 with centromeres, and cohesin loading. We propose that cohesin triggers its own loading by enabling Scc2/4 to connect with chromosomal landmarks, which at centromeres are specified by the Ctf19 complex. Overall, our findings provide a paradigm for the spatial and temporal control of cohesin loading.
Copyright © 2013 Elsevier Ltd. All rights reserved.
Figures
References
-
- Haering C.H., Löwe J., Hochwagen A., Nasmyth K. Molecular architecture of SMC proteins and the yeast cohesin complex. Mol. Cell. 2002;9:773–788. - PubMed
-
- Bernard P., Drogat J., Maure J.-F., Dheur S., Vaur S., Genier S., Javerzat J.-P. A screen for cohesion mutants uncovers Ssl3, the fission yeast counterpart of the cohesin loading factor Scc4. Curr. Biol. 2006;16:875–881. - PubMed
-
- Ciosk R., Shirayama M., Shevchenko A., Tanaka T., Toth A., Shevchenko A., Nasmyth K. Cohesin’s binding to chromosomes depends on a separate complex consisting of Scc2 and Scc4 proteins. Mol. Cell. 2000;5:243–254. - PubMed
-
- Lightfoot J., Testori S., Barroso C., Martinez-Perez E. Loading of meiotic cohesin by SCC-2 is required for early processing of DSBs and for the DNA damage checkpoint. Curr. Biol. 2011;21:1421–1430. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
Research Materials
