

Analysis by liquid chromatography-mass spectrometry of sterols and oxysterols in brain of the newborn Dhcr7(Δ3-5/T93M) mouse: a model of Smith-Lemli-Opitz syndrome
- PMID: 23500538
- PMCID: PMC4004445
- DOI: 10.1016/j.bcp.2013.03.003
Analysis by liquid chromatography-mass spectrometry of sterols and oxysterols in brain of the newborn Dhcr7(Δ3-5/T93M) mouse: a model of Smith-Lemli-Opitz syndrome
Abstract
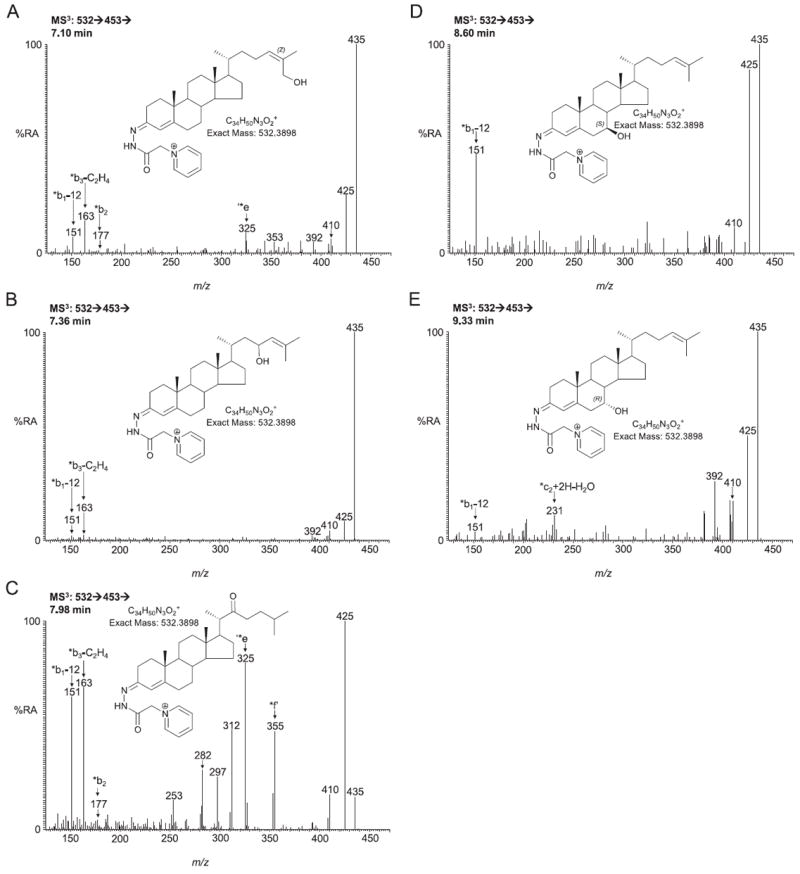
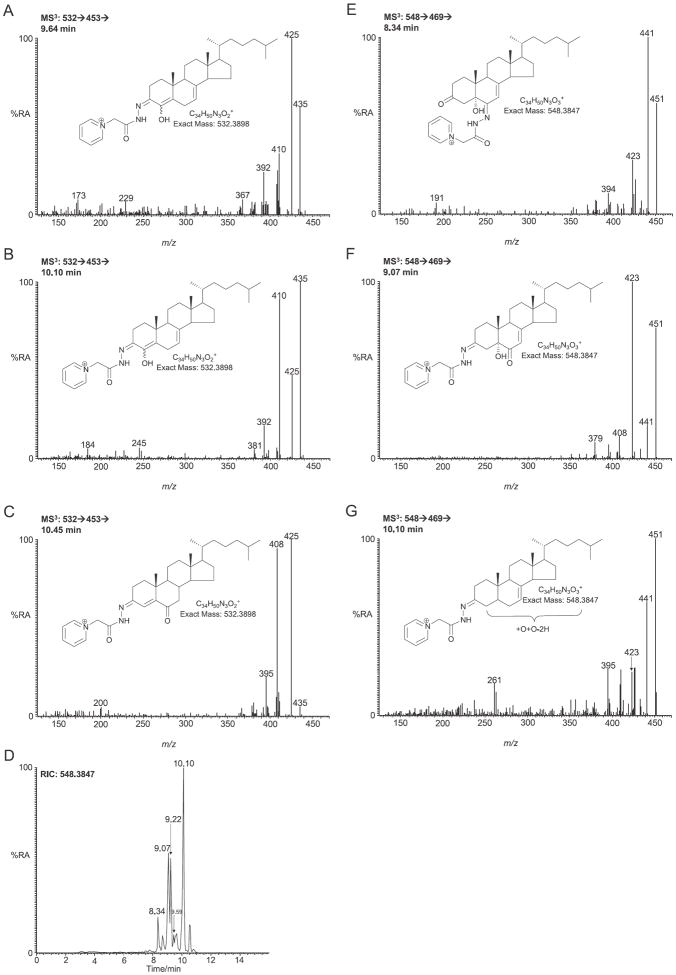
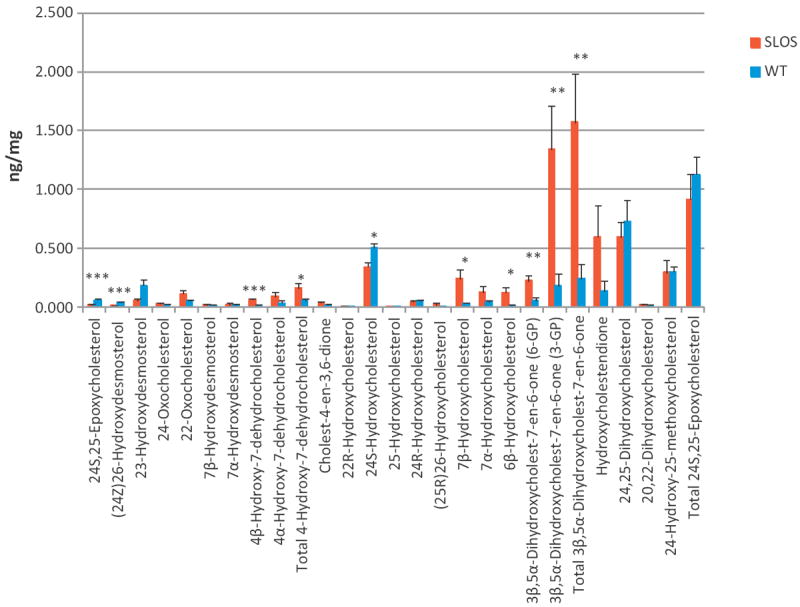
In this study the sterol and oxysterol profile of newborn brain from the Dhcr7(Δ3-5/T93M) mouse model of Smith-Lemli-Opitz syndrome (SLOS) has been investigated. This is a viable mouse model which is compound heterozygous containing one null allele and one T93M mutation on Dhcr7. We find the SLOS mouse has reduced levels of cholesterol and desmosterol and increased levels of 7- and 8-dehydrocholesterol and of 7- and 8-dehydrodesmosterol in brain compared to the wild type. The profile of enzymatically formed oxysterols in the SLOS mouse resembles that in the wild type but the level of 24S-hydroxycholesterol, the dominating cholesterol metabolite, is reduced in a similar proportion to that of cholesterol. A number of oxysterols abundant in the SLOS mouse are probably derived from 7-dehydrocholesterol, however, the mechanism of their formation is unclear.
Copyright © 2013 Elsevier Inc. All rights reserved.
Conflict of interest statement
The authors have no conflicting financial interests.
Figures
Similar articles
-
Development and characterization of a hypomorphic Smith-Lemli-Opitz syndrome mouse model and efficacy of simvastatin therapy.Hum Mol Genet. 2006 Mar 15;15(6):839-51. doi: 10.1093/hmg/ddl003. Epub 2006 Jan 30. Hum Mol Genet. 2006. PMID: 16446309
-
Hydroxyzine Effects on Post-Lanosterol Biosynthesis in Smith-Lemli-Opitz Syndrome (SLOS) Models.Biomolecules. 2025 Apr 10;15(4):562. doi: 10.3390/biom15040562. Biomolecules. 2025. PMID: 40305315 Free PMC article.
-
Antioxidant supplementation ameliorates molecular deficits in Smith-Lemli-Opitz syndrome.Biol Psychiatry. 2014 Feb 1;75(3):215-22. doi: 10.1016/j.biopsych.2013.06.013. Epub 2013 Jul 26. Biol Psychiatry. 2014. PMID: 23896203 Free PMC article.
-
Recent insights into the Smith-Lemli-Opitz syndrome.Clin Genet. 2005 Nov;68(5):383-91. doi: 10.1111/j.1399-0004.2005.00515.x. Clin Genet. 2005. PMID: 16207203 Free PMC article. Review.
-
Smith-Lemli-Opitz syndrome: pathogenesis, diagnosis and management.Eur J Hum Genet. 2008 May;16(5):535-41. doi: 10.1038/ejhg.2008.10. Epub 2008 Feb 20. Eur J Hum Genet. 2008. PMID: 18285838 Review.
Cited by
-
Oxidative stress, serotonergic changes and decreased ultrasonic vocalizations in a mouse model of Smith-Lemli-Opitz syndrome.Genes Brain Behav. 2017 Jul;16(6):619-626. doi: 10.1111/gbb.12376. Epub 2017 Mar 15. Genes Brain Behav. 2017. PMID: 28220990 Free PMC article.
-
The drug release of PLGA-based nanoparticles and their application in treatment of gastrointestinal cancers.Heliyon. 2024 Sep 19;10(18):e38165. doi: 10.1016/j.heliyon.2024.e38165. eCollection 2024 Sep 30. Heliyon. 2024. PMID: 39364250 Free PMC article. Review.
-
7-Dehydrocholesterol Encapsulated Polymeric Nanoparticles As a Radiation-Responsive Sensitizer for Enhancing Radiation Therapy.Small. 2022 Apr;18(17):e2200710. doi: 10.1002/smll.202200710. Epub 2022 Mar 18. Small. 2022. PMID: 35304816 Free PMC article.
-
Sterols and oxysterols in plasma from Smith-Lemli-Opitz syndrome patients.J Steroid Biochem Mol Biol. 2017 May;169:77-87. doi: 10.1016/j.jsbmb.2016.03.018. Epub 2016 Mar 11. J Steroid Biochem Mol Biol. 2017. PMID: 26976653 Free PMC article.
-
Pathogenesis, Epidemiology, Diagnosis and Clinical Aspects of Smith-Lemli-Opitz Syndrome.Expert Opin Orphan Drugs. 2015 Mar;3(3):267-280. doi: 10.1517/21678707.2015.1014472. Expert Opin Orphan Drugs. 2015. PMID: 25734025 Free PMC article.
References
-
- Kelley RI, Herman GE. Inborn errors of sterol biosynthesis. Annu Rev Genomics Hum Genet. 2001;2:299–341. - PubMed
-
- Björkhem I, Starck L, Andersson U, Lütjohann D, von BS, Pikuleva I, et al. Oxysterols in the circulation of patients with the Smith–Lemli–Opitz syndrome: abnormal levels of 24S- and 27-hydroxycholesterol. J Lipid Res. 2001;42:366–71. - PubMed
-
- Kelley RI. Diagnosis of Smith–Lemli–Opitz syndrome by gas chromatography/mass spectrometry of 7-dehydrocholesterol in plasma, amniotic fluid and cultured skin fibroblasts. Clin Chim Acta. 1995;236:45–58. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
- BB/C515771/2/BB_/Biotechnology and Biological Sciences Research Council/United Kingdom
- HD053036/HD/NICHD NIH HHS/United States
- R01 HD053036/HD/NICHD NIH HHS/United States
- BBH0010181/BB_/Biotechnology and Biological Sciences Research Council/United Kingdom
- BBC5157712/BB_/Biotechnology and Biological Sciences Research Council/United Kingdom
- BBI0017351/BB_/Biotechnology and Biological Sciences Research Council/United Kingdom
- BB/C511356/1/BB_/Biotechnology and Biological Sciences Research Council/United Kingdom
- BBC5113561/BB_/Biotechnology and Biological Sciences Research Council/United Kingdom
- BB/H001018/1/BB_/Biotechnology and Biological Sciences Research Council/United Kingdom
LinkOut - more resources
Full Text Sources
Other Literature Sources
Miscellaneous
