

The exoribonuclease Dis3L2 defines a novel eukaryotic RNA degradation pathway
- PMID: 23503588
- PMCID: PMC3981172
- DOI: 10.1038/emboj.2013.63
The exoribonuclease Dis3L2 defines a novel eukaryotic RNA degradation pathway
Abstract
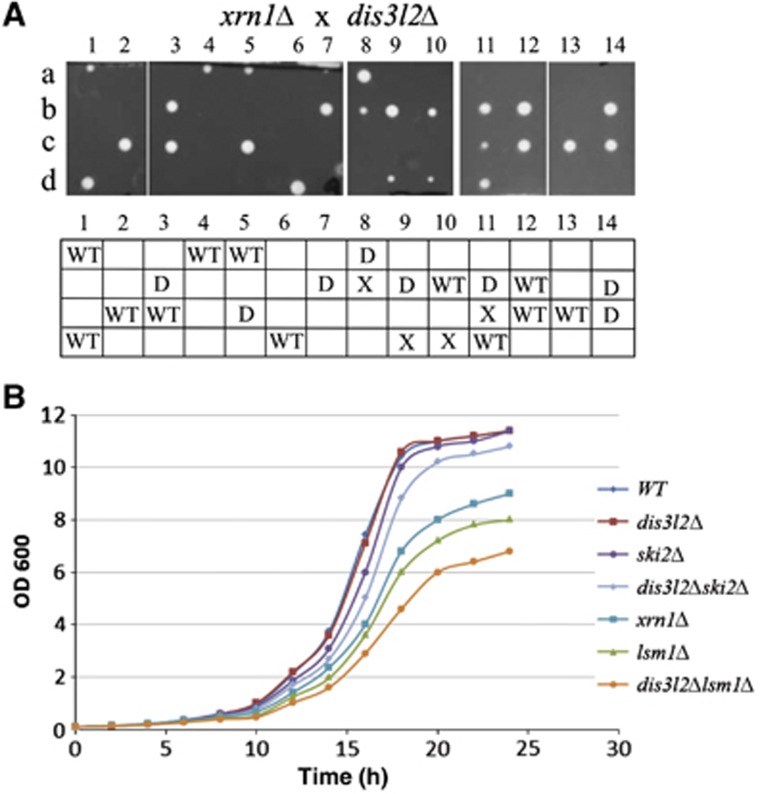
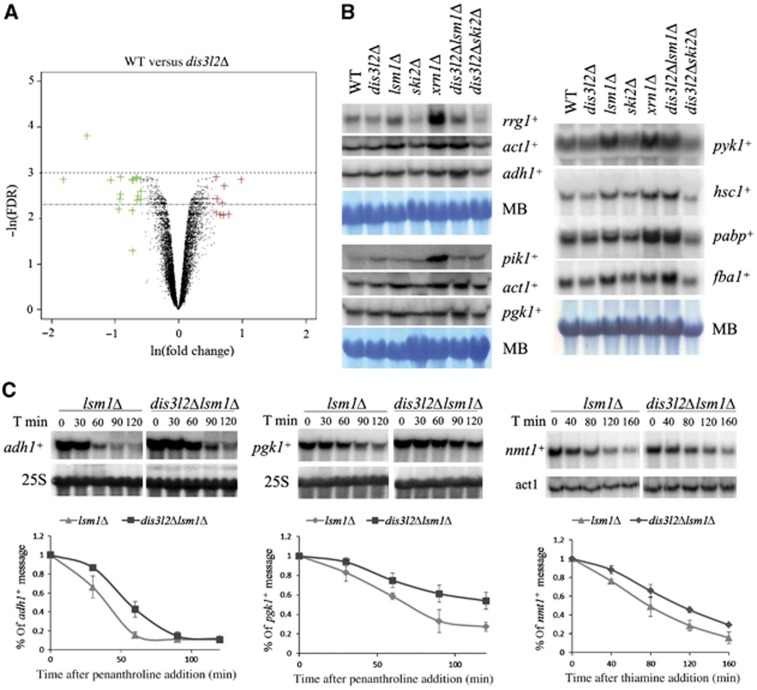
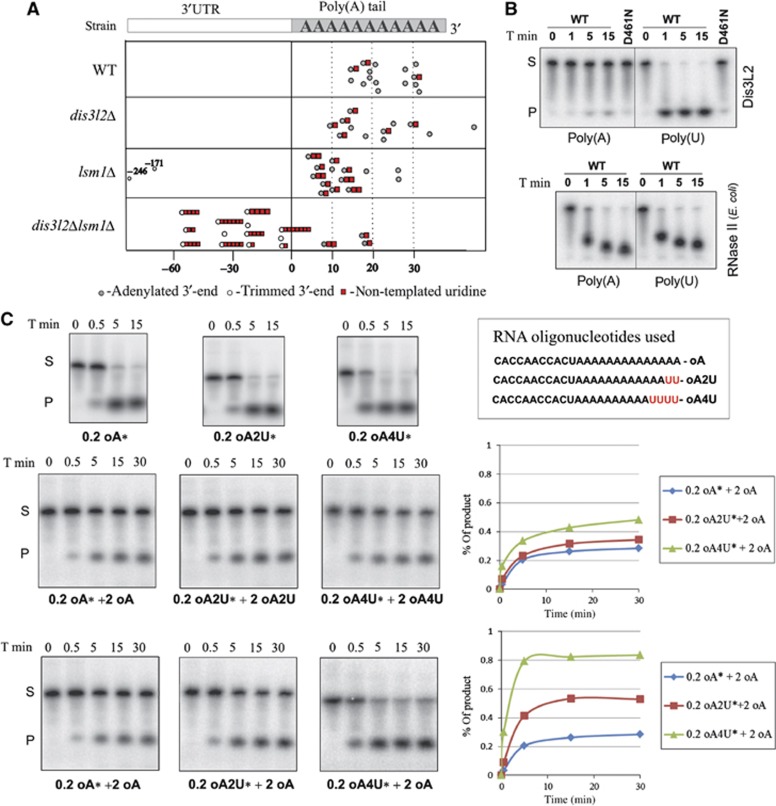
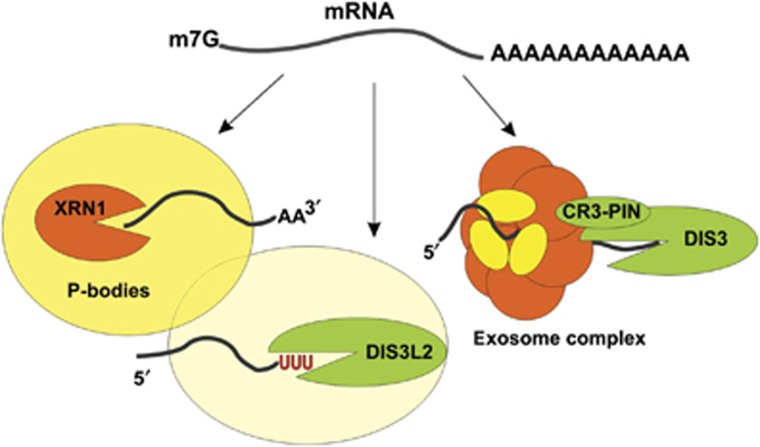
The final step of cytoplasmic mRNA degradation proceeds in either a 5'-3' direction catalysed by Xrn1 or in a 3'-5' direction catalysed by the exosome. Dis3/Rrp44, an RNase II family protein, is the catalytic subunit of the exosome. In humans, there are three paralogues of this enzyme: DIS3, DIS3L, and DIS3L2. In this work, we identified a novel Schizosaccharomyces pombe exonuclease belonging to the conserved family of human DIS3L2 and plant SOV. Dis3L2 does not interact with the exosome components and localizes in the cytoplasm and in cytoplasmic foci, which are docked to P-bodies. Deletion of dis3l2(+) is synthetically lethal with xrn1Δ, while deletion of dis3l2(+) in an lsm1Δ background results in the accumulation of transcripts and slower mRNA degradation rates. Accumulated transcripts show enhanced uridylation and in vitro Dis3L2 displays a preference for uridylated substrates. Altogether, our results suggest that in S. pombe, and possibly in most other eukaryotes, Dis3L2 is an important factor in mRNA degradation. Therefore, this novel 3'-5' RNA decay pathway represents an alternative to degradation by Xrn1 and the exosome.
Conflict of interest statement
The authors declare that they have no conflict of interest.
Figures
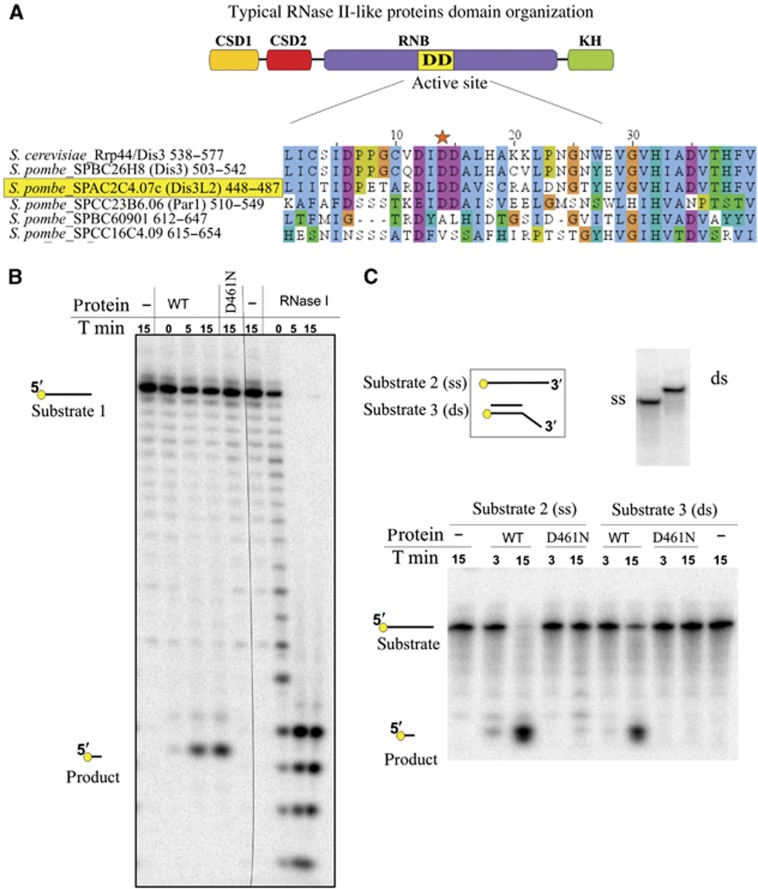
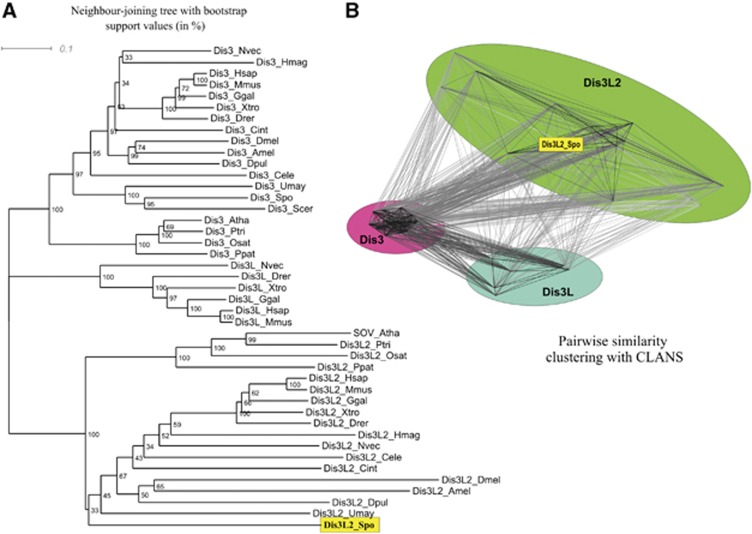
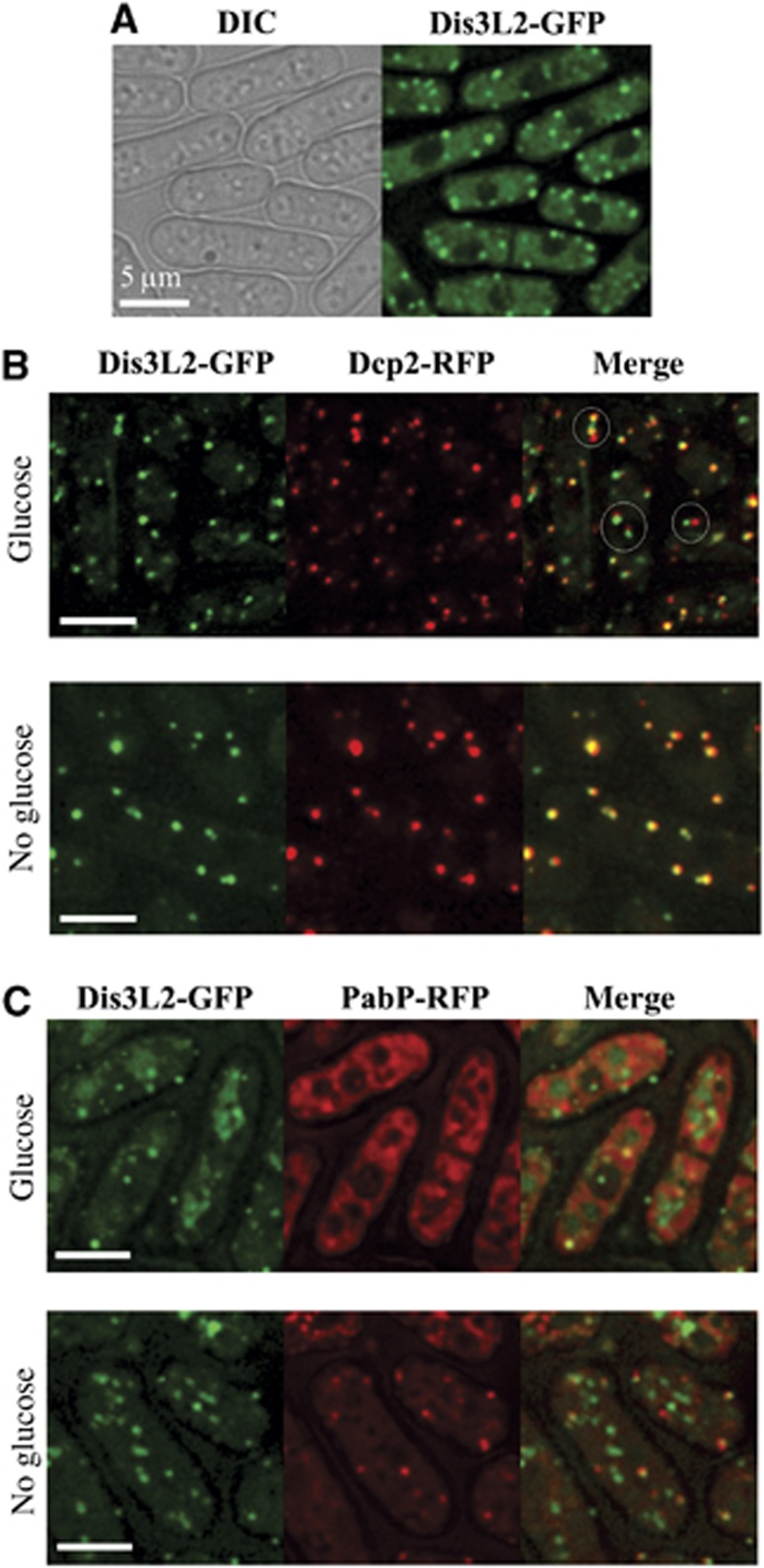
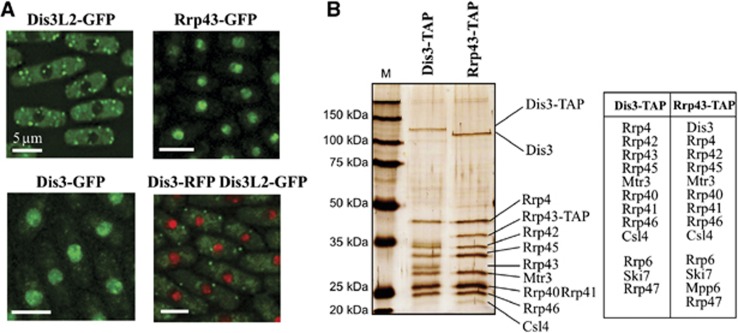
Comment in
-
A DIStinctively novel exoribonuclease that really likes U.EMBO J. 2013 Jul 3;32(13):1799-801. doi: 10.1038/emboj.2013.136. Epub 2013 Jun 11. EMBO J. 2013. PMID: 23756464 Free PMC article.
References
-
- Alexander RD, Barrass JD, Dichtl B, Kos M, Obtulowicz T, Robert MC, Koper M, Karkusiewicz I, Mariconti L, Tollervey D, Kufel J, Bertrand E, Beggs JD (2010) RiboSys, a high-resolution, quantitative approach to measure the in vivo kinetics of pre-mRNA splicing and 3′-end processing in Saccharomyces cerevisiae. RNA 16: 2570–2580 - PMC - PubMed
-
- Amblar M, Arraiano CM (2005) A single mutation in Escherichia coli ribonuclease II inactivates the enzyme without affecting RNA binding. FEBS J 272: 363–374 - PubMed
-
- Anderson P, Kedersha N (2008) Stress granules: the Tao of RNA triage. Trends Biochem Sci 33: 141–150 - PubMed
-
- Astuti D, Morris MR, Cooper WN, Staals RH, Wake NC, Fews GA, Gill H, Gentle D, Shuib S, Ricketts CJ, Cole T, van Essen AJ, van Lingen RA, Neri G, Opitz JM, Rump P, Stolte-Dijkstra I, Muller F, Pruijn GJ, Latif F et al. (2012) Germline mutations in DIS3L2 cause the Perlman syndrome of overgrowth and Wilms tumor susceptibility. Nat Genet 44: 277–284 - PubMed
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
