

Disruption of SMIM1 causes the Vel- blood type
- PMID: 23505126
- PMCID: PMC3662317
- DOI: 10.1002/emmm.201302466
Disruption of SMIM1 causes the Vel- blood type
Abstract
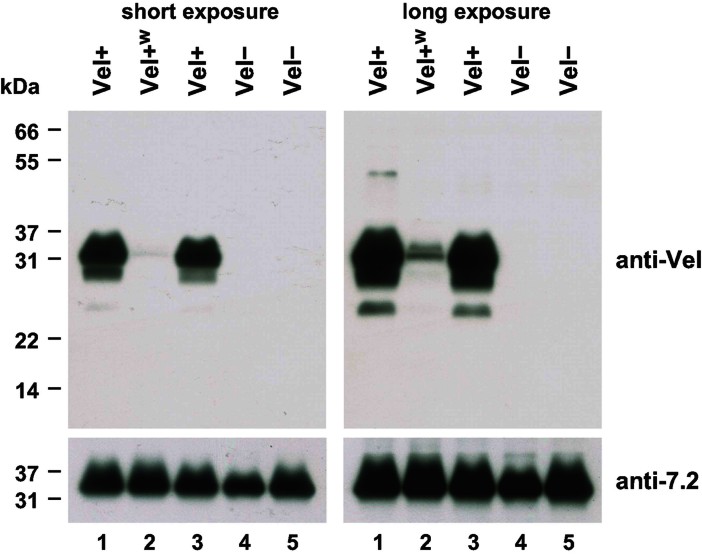
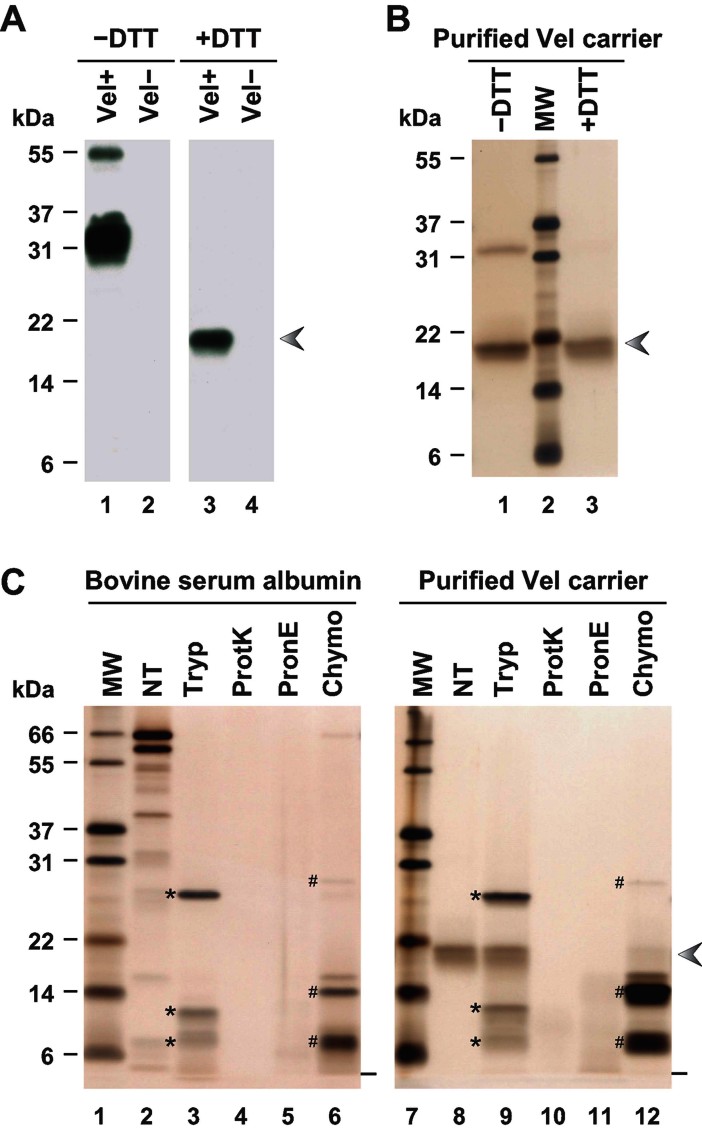
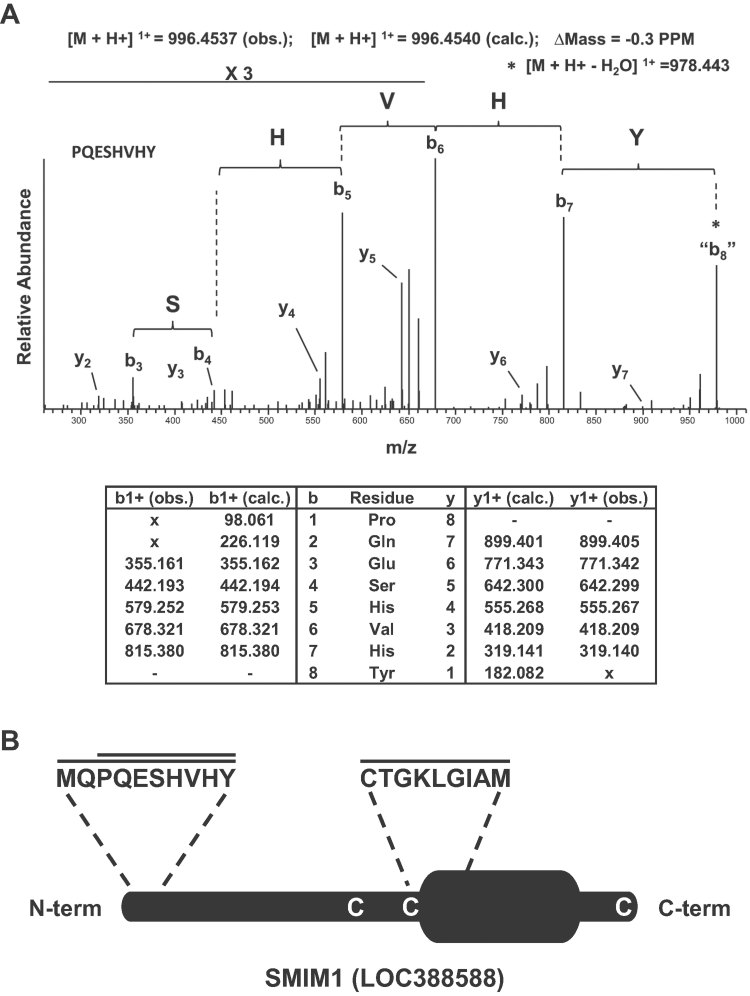
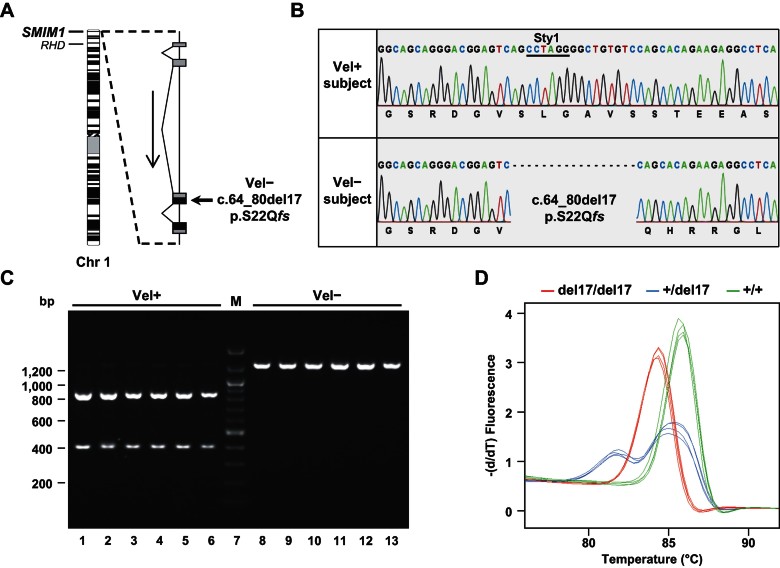
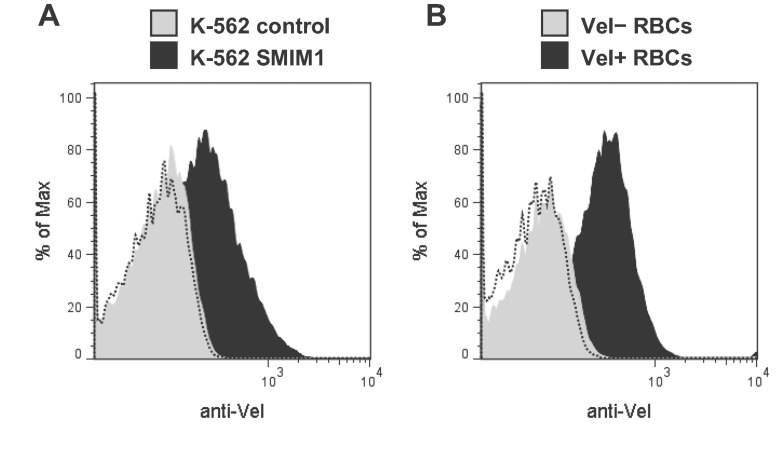
Here, we report the biochemical and genetic basis of the Vel blood group antigen, which has been a vexing mystery for decades, especially as anti-Vel regularly causes severe haemolytic transfusion reactions. The protein carrying the Vel blood group antigen was biochemically purified from red blood cell membranes. Mass spectrometry-based de novo peptide sequencing identified this protein to be small integral membrane protein 1 (SMIM1), a previously uncharacterized single-pass membrane protein. Expression of SMIM1 cDNA in Vel- cultured cells generated anti-Vel cell surface reactivity, confirming that SMIM1 encoded the Vel blood group antigen. A cohort of 70 Vel- individuals was found to be uniformly homozygous for a 17 nucleotide deletion in the coding sequence of SMIM1. The genetic homogeneity of the Vel- blood type, likely having a common origin, facilitated the development of two highly specific DNA-based tests for rapid Vel genotyping, which can be easily integrated into blood group genotyping platforms. These results answer a 60-year-old riddle and provide tools of immediate assistance to all clinicians involved in the care of Vel- patients.
Copyright © 2013 The Authors. Published by John Wiley and Sons, Ltd on behalf of EMBO.
Figures
References
-
- Alfred BM, Stout TD, Lee M, Birkbeck J, Petrakis NL. Blood groups, phosphoglucomutase, and cerumen types of the Anaham (Chilcotin) Indians. Am J Phys Anthropol. 1970;32:329–337. - PubMed
-
- Allen FH, Jr, Corcoran PA. Blood groups of the Penobscot Indians. Am J Phys Anthropol. 1960;18:109–114. - PubMed
-
- Altschul SF, Gish W, Miller W, Myers EW, Lipman DJ. Basic local alignment search tool. J Mol Biol. 1990;215:403–410. - PubMed
-
- Ballif BA, Cao Z, Schwartz D, Carraway KL, III, Gygi SP. Identification of 14-3-3epsilon substrates from embryonic murine brain. J Proteome Res. 2006;5:2372–2379. - PubMed
-
- Ballif BA, Carey GR, Sunyaev SR, Gygi SP. Large-scale identification and evolution indexing of tyrosine phosphorylation sites from murine brain. J Proteome Res. 2008;7:311–318. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
Research Materials
