

A unifying mechanism for cancer cell death through ion channel activation by HAMLET
- PMID: 23505537
- PMCID: PMC3591364
- DOI: 10.1371/journal.pone.0058578
A unifying mechanism for cancer cell death through ion channel activation by HAMLET
Abstract
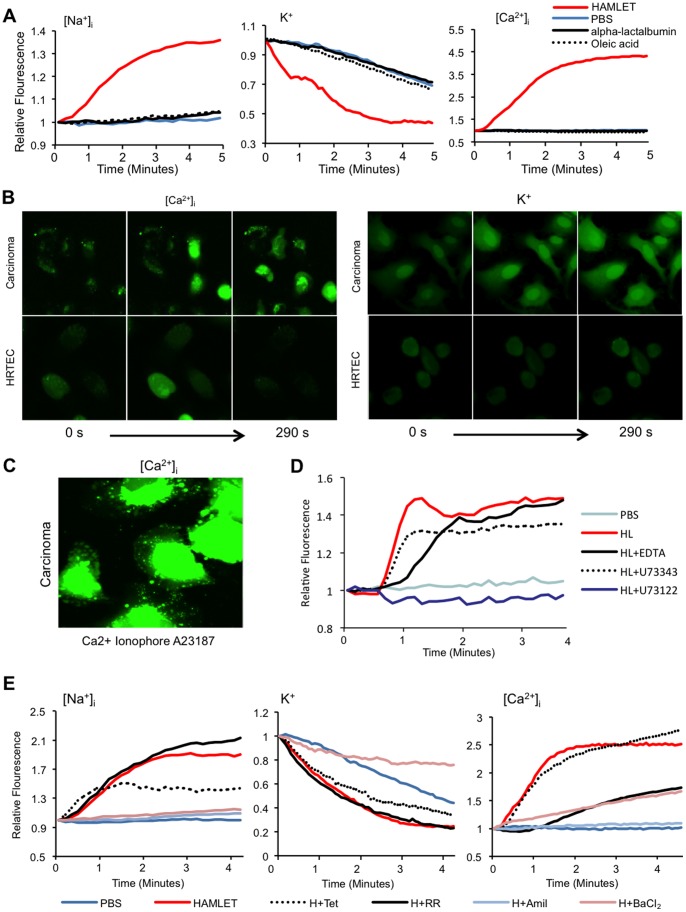
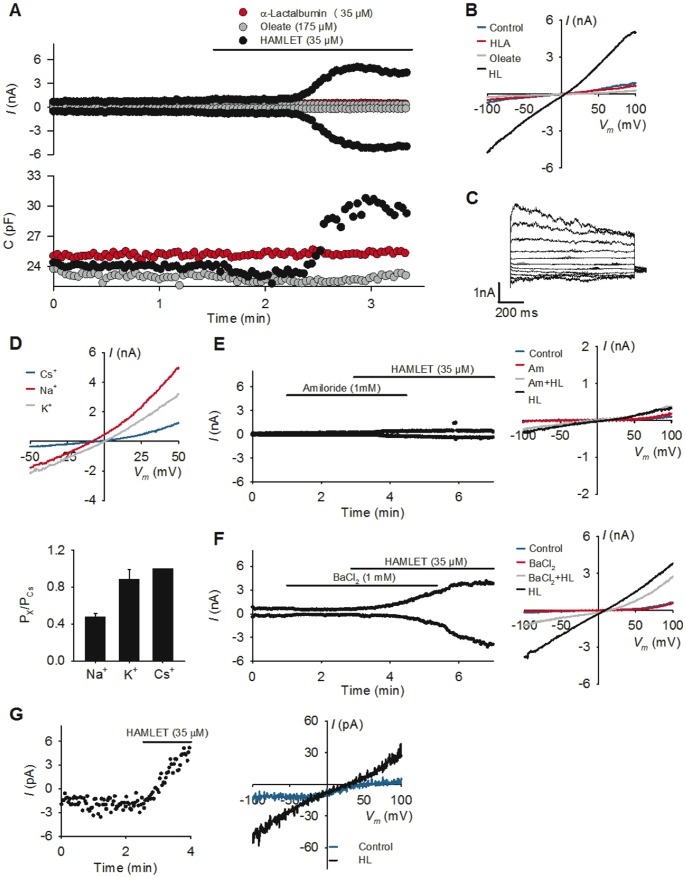
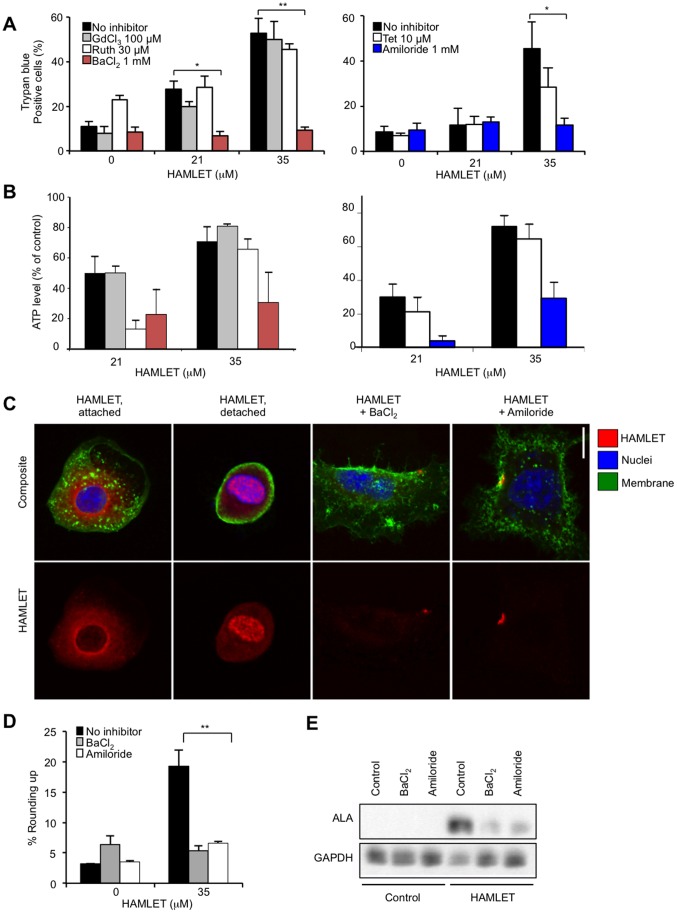
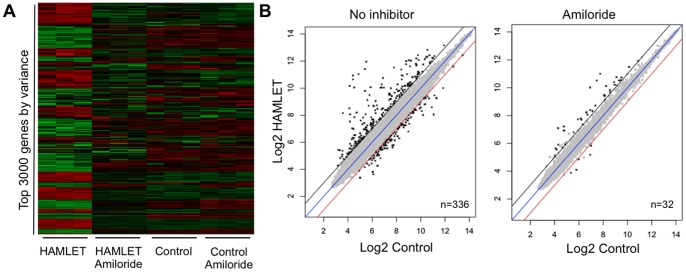
Ion channels and ion fluxes control many aspects of tissue homeostasis. During oncogenic transformation, critical ion channel functions may be perturbed but conserved tumor specific ion fluxes remain to be defined. Here we used the tumoricidal protein-lipid complex HAMLET as a probe to identify ion fluxes involved in tumor cell death. We show that HAMLET activates a non-selective cation current, which reached a magnitude of 2.74±0.88 nA within 1.43±0.13 min from HAMLET application. Rapid ion fluxes were essential for HAMLET-induced carcinoma cell death as inhibitors (amiloride, BaCl2), preventing the changes in free cellular Na(+) and K(+) concentrations also prevented essential steps accompanying carcinoma cell death, including changes in morphology, uptake, global transcription, and MAP kinase activation. Through global transcriptional analysis and phosphorylation arrays, a strong ion flux dependent p38 MAPK response was detected and inhibition of p38 signaling delayed HAMLET-induced death. Healthy, differentiated cells were resistant to HAMLET challenge, which was accompanied by innate immunity rather than p38-activation. The results suggest, for the first time, a unifying mechanism for the initiation of HAMLET's broad and rapid lethal effect on tumor cells. These findings are particularly significant in view of HAMLET's documented therapeutic efficacy in human studies and animal models. The results also suggest that HAMLET offers a two-tiered therapeutic approach, killing cancer cells while stimulating an innate immune response in surrounding healthy tissues.
Conflict of interest statement
Figures
Similar articles
-
HAMLET binding to α-actinin facilitates tumor cell detachment.PLoS One. 2011 Mar 8;6(3):e17179. doi: 10.1371/journal.pone.0017179. PLoS One. 2011. PMID: 21408150 Free PMC article.
-
Conserved features of cancer cells define their sensitivity to HAMLET-induced death; c-Myc and glycolysis.Oncogene. 2011 Dec 1;30(48):4765-79. doi: 10.1038/onc.2011.196. Epub 2011 Jun 6. Oncogene. 2011. PMID: 21643007 Free PMC article.
-
Targeting of nucleotide-binding proteins by HAMLET--a conserved tumor cell death mechanism.Oncogene. 2016 Feb 18;35(7):897-907. doi: 10.1038/onc.2015.144. Epub 2015 Jun 1. Oncogene. 2016. PMID: 26028028
-
HAMLET: functional properties and therapeutic potential.Future Oncol. 2012 Oct;8(10):1301-13. doi: 10.2217/fon.12.122. Future Oncol. 2012. PMID: 23130929 Review.
-
HAMLET - A protein-lipid complex with broad tumoricidal activity.Biochem Biophys Res Commun. 2017 Jan 15;482(3):454-458. doi: 10.1016/j.bbrc.2016.10.092. Epub 2017 Feb 3. Biochem Biophys Res Commun. 2017. PMID: 28212731 Review.
Cited by
-
A Characeae Cells Plasma Membrane as a Model for Selection of Bioactive Compounds and Drugs: Interaction of HAMLET-Like Complexes with Ion Channels of Chara corallina Cells Plasmalemma.J Membr Biol. 2016 Dec;249(6):801-811. doi: 10.1007/s00232-016-9930-1. Epub 2016 Sep 16. J Membr Biol. 2016. PMID: 27638176
-
α-Lactalbumin, Amazing Calcium-Binding Protein.Biomolecules. 2020 Aug 20;10(9):1210. doi: 10.3390/biom10091210. Biomolecules. 2020. PMID: 32825311 Free PMC article. Review.
-
Protein-lipid complexes: molecular structure, current scenarios and mechanisms of cytotoxicity.RSC Adv. 2019 Nov 13;9(63):36890-36906. doi: 10.1039/c9ra07127j. eCollection 2019 Nov 11. RSC Adv. 2019. PMID: 35539089 Free PMC article. Review.
-
Protein receptor-independent plasma membrane remodeling by HAMLET: a tumoricidal protein-lipid complex.Sci Rep. 2015 Nov 12;5:16432. doi: 10.1038/srep16432. Sci Rep. 2015. PMID: 26561036 Free PMC article.
-
Bladder cancer therapy using a conformationally fluid tumoricidal peptide complex.Nat Commun. 2021 Jun 8;12(1):3427. doi: 10.1038/s41467-021-23748-y. Nat Commun. 2021. PMID: 34103518 Free PMC article. Clinical Trial.
References
-
- Martinac B (2004) Mechanosensitive ion channels: molecules of mechanotransduction. Journal of cell science 117: 2449–2460. - PubMed
-
- Nilius B, Owsianik G, Voets T, Peters JA (2007) Transient receptor potential cation channels in disease. Physiol Rev 87: 165–217. - PubMed
-
- Pedersen SF, Nilius B (2007) Transient receptor potential channels in mechanosensing and cell volume regulation. Methods Enzymol 428: 183–207. - PubMed
-
- Arcangeli A (2011) Ion channels and transporters in cancer. 3. Ion channels in the tumor cell-microenvironment cross talk. Am J Physiol Cell Physiol 301: C762–771. - PubMed
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
