

The protein translocation systems in plants - composition and variability on the example of Solanum lycopersicum
- PMID: 23506162
- PMCID: PMC3610429
- DOI: 10.1186/1471-2164-14-189
The protein translocation systems in plants - composition and variability on the example of Solanum lycopersicum
Abstract
Background: Protein translocation across membranes is a central process in all cells. In the past decades the molecular composition of the translocation systems in the membranes of the endoplasmic reticulum, peroxisomes, mitochondria and chloroplasts have been established based on the analysis of model organisms. Today, these results have to be transferred to other plant species. We bioinformatically determined the inventory of putative translocation factors in tomato (Solanum lycopersicum) by orthologue search and domain architecture analyses. In addition, we investigated the diversity of such systems by comparing our findings to the model organisms Saccharomyces cerevisiae, Arabidopsis thaliana and 12 other plant species.
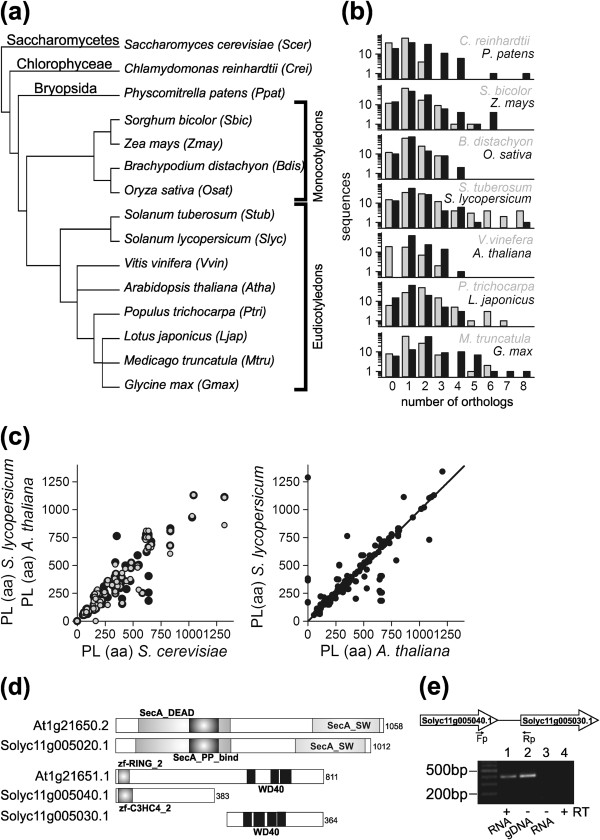
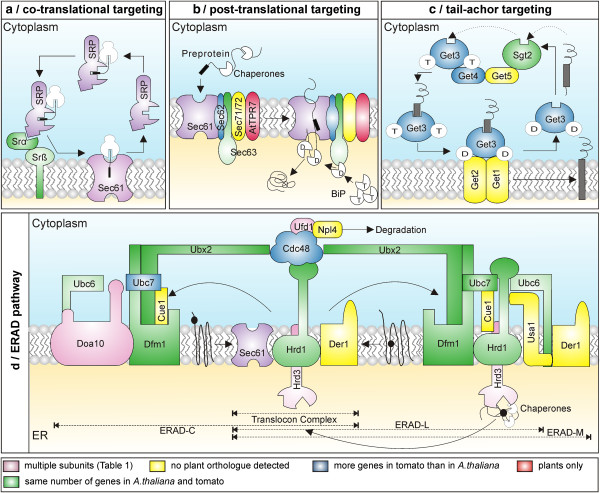
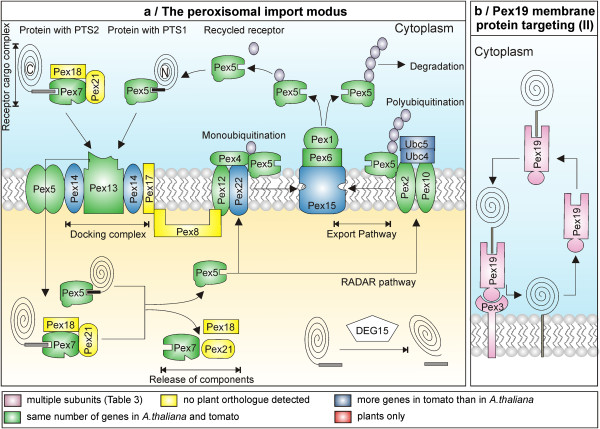
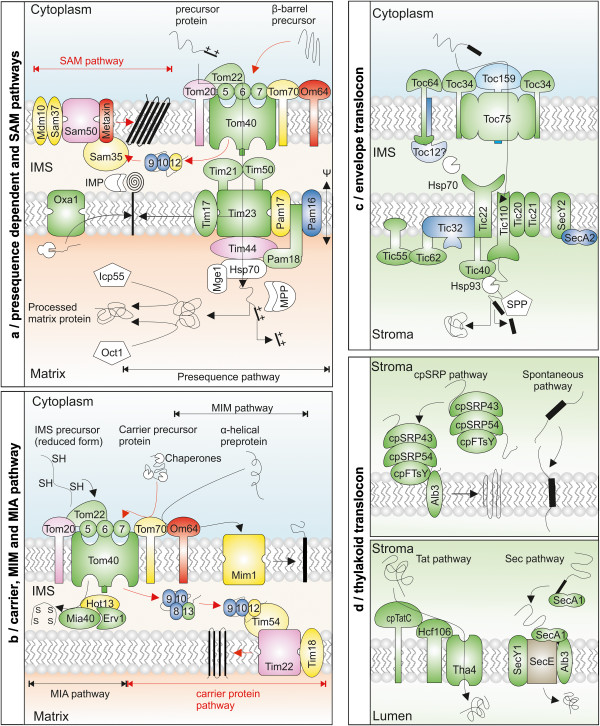
Results: The literature search end up in a total of 130 translocation components in yeast and A. thaliana, which are either experimentally confirmed or homologous to experimentally confirmed factors. From our bioinformatic analysis (PGAP and OrthoMCL), we identified (co-)orthologues in plants, which in combination yielded 148 and 143 orthologues in A. thaliana and S. lycopersicum, respectively. Interestingly, we traced 82% overlap in findings from both approaches though we did not find any orthologues for 27% of the factors by either procedure. In turn, 29% of the factors displayed the presence of more than one (co-)orthologue in tomato. Moreover, our analysis revealed that the genomic composition of the translocation machineries in the bryophyte Physcomitrella patens resemble more to higher plants than to single celled green algae. The monocots (Z. mays and O. sativa) follow more or less a similar conservation pattern for encoding the translocon components. In contrast, a diverse pattern was observed in different eudicots.
Conclusions: The orthologue search shows in most cases a clear conservation of components of the translocation pathways/machineries. Only the Get-dependent integration of tail-anchored proteins seems to be distinct. Further, the complexity of the translocation pathway in terms of existing orthologues seems to vary among plant species. This might be the consequence of palaeoploidisation during evolution in plants; lineage specific whole genome duplications in Arabidopsis thaliana and triplications in Solanum lycopersicum.
Figures
Similar articles
-
The complexity of vesicle transport factors in plants examined by orthology search.PLoS One. 2014 May 20;9(5):e97745. doi: 10.1371/journal.pone.0097745. eCollection 2014. PLoS One. 2014. PMID: 24844592 Free PMC article.
-
The intracellular distribution of the components of the GET system in vascular plants.Biochim Biophys Acta Mol Cell Res. 2019 Oct;1866(10):1650-1662. doi: 10.1016/j.bbamcr.2019.06.012. Epub 2019 Jun 21. Biochim Biophys Acta Mol Cell Res. 2019. PMID: 31233800
-
Survey of Genes Involved in Biosynthesis, Transport, and Signaling of Phytohormones with Focus on Solanum lycopersicum.Bioinform Biol Insights. 2016 Sep 26;10:185-207. doi: 10.4137/BBI.S38425. eCollection 2016. Bioinform Biol Insights. 2016. PMID: 27695302 Free PMC article.
-
Translating the Arabidopsis thaliana Peroxisome Proteome Insights to Solanum lycopersicum: Consensus Versus Diversity.Front Cell Dev Biol. 2022 Jul 13;10:909604. doi: 10.3389/fcell.2022.909604. eCollection 2022. Front Cell Dev Biol. 2022. PMID: 35912119 Free PMC article. Review.
-
Molecular basis of plant architecture.Annu Rev Plant Biol. 2008;59:253-79. doi: 10.1146/annurev.arplant.59.032607.092902. Annu Rev Plant Biol. 2008. PMID: 18444901 Review.
Cited by
-
Comparative analysis of putative orthologues of mitochondrial import motor subunit: Pam18 and Pam16 in plants.PLoS One. 2013 Oct 23;8(10):e78400. doi: 10.1371/journal.pone.0078400. eCollection 2013. PLoS One. 2013. PMID: 24194927 Free PMC article.
-
The bs5 allele of the susceptibility gene Bs5 of pepper (Capsicum annuum L.) encoding a natural deletion variant of a CYSTM protein conditions resistance to bacterial spot disease caused by Xanthomonas species.Theor Appl Genet. 2023 Mar 21;136(3):64. doi: 10.1007/s00122-023-04340-y. Theor Appl Genet. 2023. PMID: 36943531 Free PMC article.
-
Protein Targeting Into the Thylakoid Membrane Through Different Pathways.Front Physiol. 2022 Jan 12;12:802057. doi: 10.3389/fphys.2021.802057. eCollection 2021. Front Physiol. 2022. PMID: 35095563 Free PMC article. Review.
-
Reliability of plastid and mitochondrial localisation prediction declines rapidly with the evolutionary distance to the training set increasing.PLoS Comput Biol. 2024 Nov 11;20(11):e1012575. doi: 10.1371/journal.pcbi.1012575. eCollection 2024 Nov. PLoS Comput Biol. 2024. PMID: 39527633 Free PMC article.
-
The complexity of vesicle transport factors in plants examined by orthology search.PLoS One. 2014 May 20;9(5):e97745. doi: 10.1371/journal.pone.0097745. eCollection 2014. PLoS One. 2014. PMID: 24844592 Free PMC article.
References
-
- Bolte K, Gruenheit N, Felsner G, Sommer MS, Maier UG, Hempel F. Making new out of old: recycling and modification of an ancient protein translocation system during eukaryotic evolution. Mechanistic comparison and phylogenetic analysis of ERAD, SELMA and the peroxisomal importomer. Bioessays. 2011;33(5):368–376. doi: 10.1002/bies.201100007. - DOI - PubMed
Publication types
MeSH terms
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
Miscellaneous
