

Ras and extracellular signal-regulated kinase signaling in thymocytes and T cells
- PMID: 23506953
- PMCID: PMC3856398
- DOI: 10.1016/j.it.2013.02.004
Ras and extracellular signal-regulated kinase signaling in thymocytes and T cells
Abstract
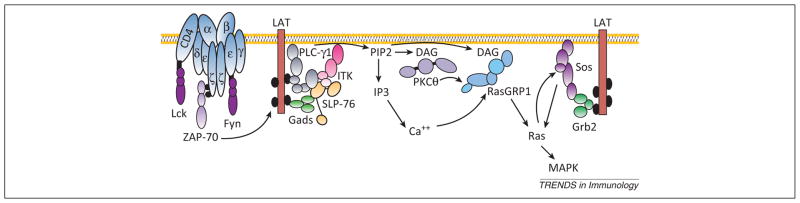
Extracellular signal-regulated kinase (ERK) activation is important for both thymocyte development and T cell function. Classically, signal transduction from the T cell antigen receptor (TCR) to ERK is thought to be regulated by signaling from Ras guanine nucleotide exchange factors (GEFs), through the small G protein Ras, to the three-tiered Raf-MAPK/ERK kinase (MEK)-ERK kinase cascade. Developing and mature T cells express four members of two RasGEF families, RasGRP1, RasGRP4, son of sevenless 1 (Sos1), and Sos2, and several models describing combined signaling from these RasGEFs have been proposed. However, recent studies suggest that existing models need revision to include both distinct and overlapping roles of multiple RasGEFs during thymocyte development and novel, Ras-independent signals to ERK that have been identified in peripheral T cells.
Published by Elsevier Ltd.
Figures


References
-
- Downward J. Targeting RAS signalling pathways in cancer therapy. Nat Rev Cancer. 2003;3:11–22. - PubMed
-
- Roberts PJ, Der CJ. Targeting the Raf-MEK-ERK mitogen-activated protein kinase cascade for the treatment of cancer. Oncogene. 2007;26:3291–3310. - PubMed
-
- Aguado E, et al. Induction of T helper type 2 immunity by a point mutation in the LAT adaptor. Science. 2002;296:2036–2040. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Research Materials
Miscellaneous
