

Transmembrane signaling and assembly of the cytochrome b6f-lipidic charge transfer complex
- PMID: 23507619
- PMCID: PMC4029431
- DOI: 10.1016/j.bbabio.2013.03.002
Transmembrane signaling and assembly of the cytochrome b6f-lipidic charge transfer complex
Abstract
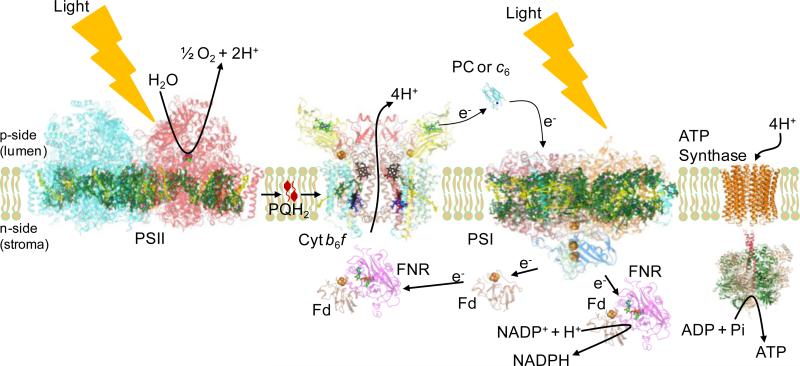
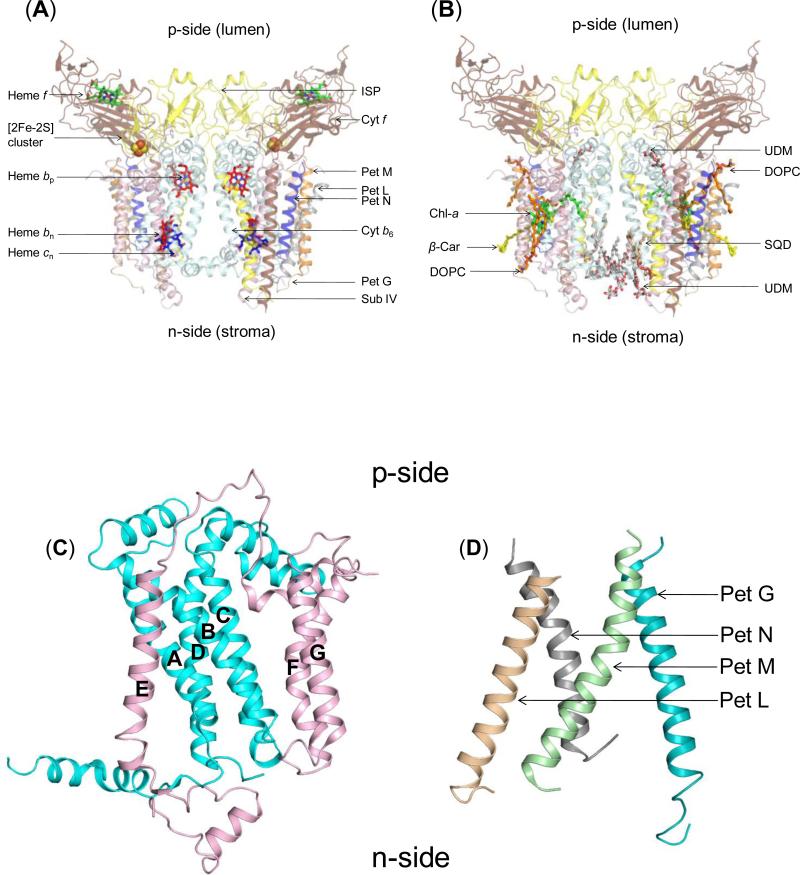
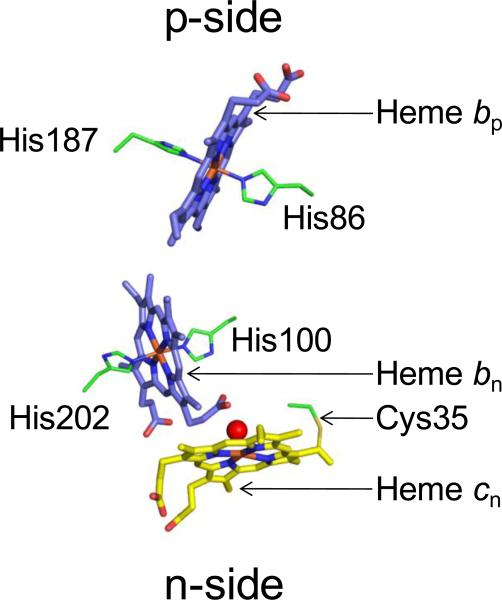
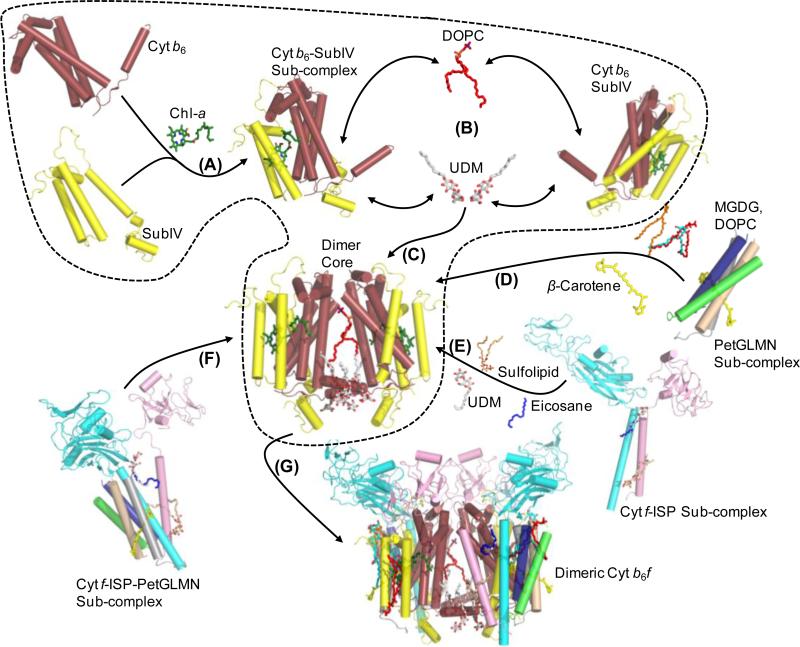
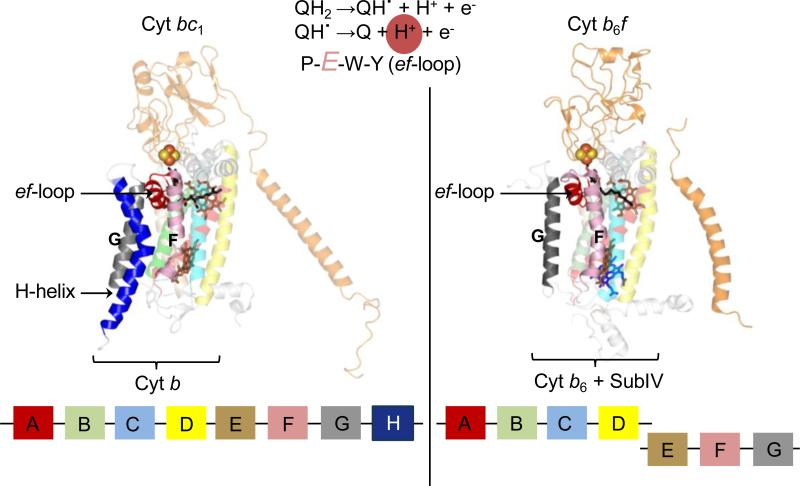
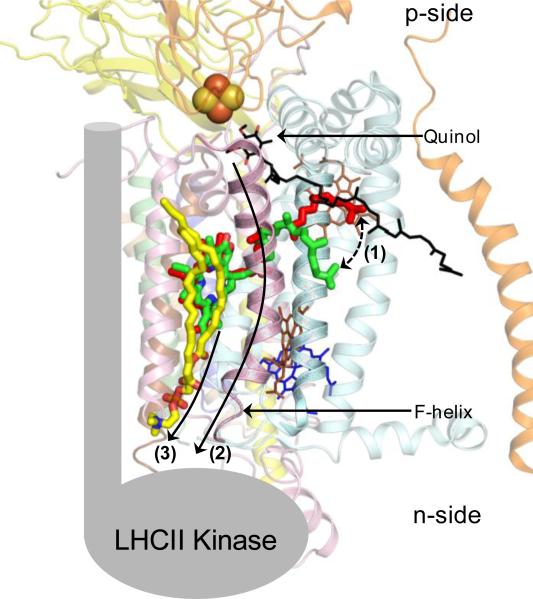
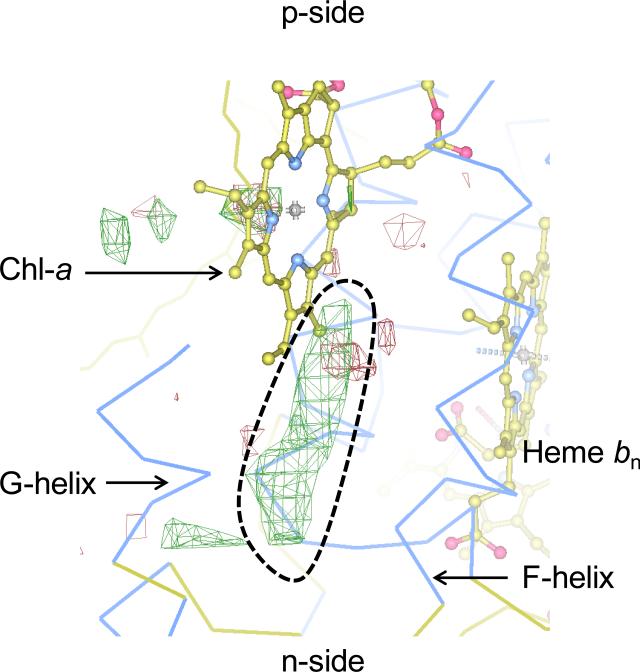
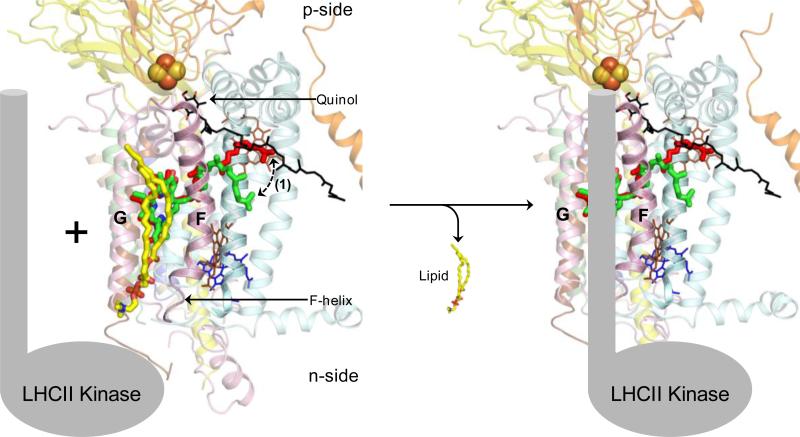
Structure-function properties of the cytochrome b6f complex are sufficiently unique compared to those of the cytochrome bc1 complex that b6f should not be considered a trivially modified bc1 complex. A unique property of the dimeric b6f complex is its involvement in transmembrane signaling associated with the p-side oxidation of plastoquinol. Structure analysis of lipid binding sites in the cyanobacterial b6f complex prepared by hydrophobic chromatography shows that the space occupied by the H transmembrane helix in the cytochrome b subunit of the bc1 complex is mostly filled by a lipid in the b6f crystal structure. It is suggested that this space can be filled by the domain of a transmembrane signaling protein. The identification of lipid sites and likely function defines the intra-membrane conserved central core of the b6f complex, consisting of the seven trans-membrane helices of the cytochrome b and subunit IV polypeptides. The other six TM helices, contributed by cytochrome f, the iron-sulfur protein, and the four peripheral single span subunits, define a peripheral less conserved domain of the complex. The distribution of conserved and non-conserved domains of each monomer of the complex, and the position and inferred function of a number of the lipids, suggests a model for the sequential assembly in the membrane of the eight subunits of the b6f complex, in which the assembly is initiated by formation of the cytochrome b6-subunit IV core sub-complex in a monomer unit. Two conformations of the unique lipidic chlorophyll a, defined in crystal structures, are described, and functions of the outlying β-carotene, a possible 'latch' in supercomplex formation, are discussed. This article is part of a Special Issue entitled: Respiratory complex III and related bc complexes.
Keywords: Assembly; Chl-a; Cyt; Cytochrome; Em7; LHC kinase; PC; PDB; PQ; PS; TMH; Transmembrane signaling; b(6)f; bc(1); chlorophyll-a; cytochrome; midpoint oxidation-reduction potential at pH7; n; p-sides, electrochemically negative and positive side of the membrane; photosystem; plastocyanin; plastoquinone; protein data bank; subIV; subunit IV of the b(6)f complex; transmembrane helix; transmembrane proton electrochemical potential gradient; Δμ̃H+; β-Car; β-carotene.
Copyright © 2013 Elsevier B.V. All rights reserved.
Figures





References
-
- Nelson N, Ben-Shem A. The complex architecture of oxygenic photosynthesis, Nature reviews. Molecular cell biology. 2004;5:971–982. - PubMed
-
- Kurisu G, Zhang H, Smith JL, Cramer WA. Structure of the cytochrome b6f complex of oxygenic photosynthesis: tuning the cavity. Science. 2003;302:1009–1014. - PubMed
-
- Stroebel D, Choquet Y, Popot JL, Picot D. An atypical haem in the cytochrome b(6)f complex. Nature. 2003;426:413–418. - PubMed
-
- Allen JF. Cytochrome b6f: structure for signalling and vectorial metabolism. Trends in plant science. 2004;9:130–137. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
