

Organization of the extracellular portion of the macrophage galactose receptor: a trimeric cluster of simple binding sites for N-acetylgalactosamine
- PMID: 23507965
- PMCID: PMC3671775
- DOI: 10.1093/glycob/cwt022
Organization of the extracellular portion of the macrophage galactose receptor: a trimeric cluster of simple binding sites for N-acetylgalactosamine
Abstract
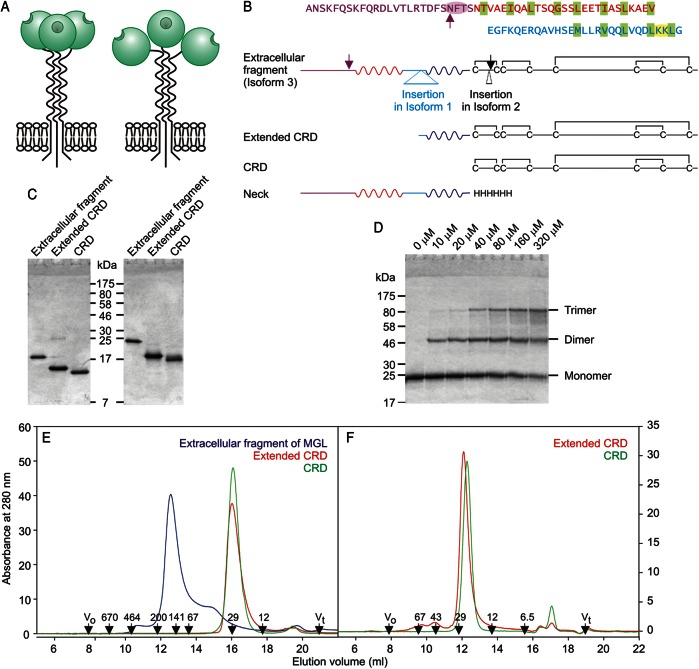
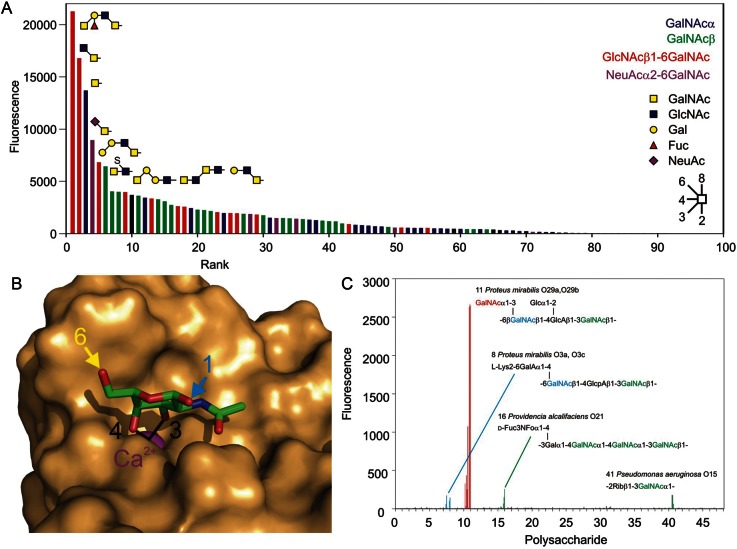
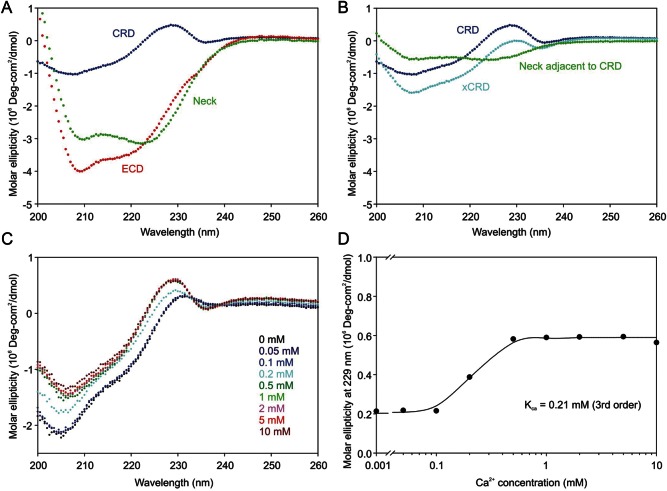
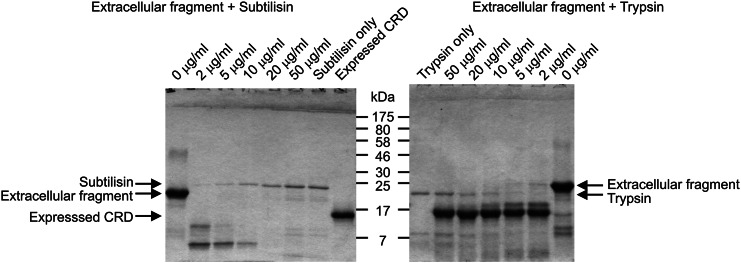
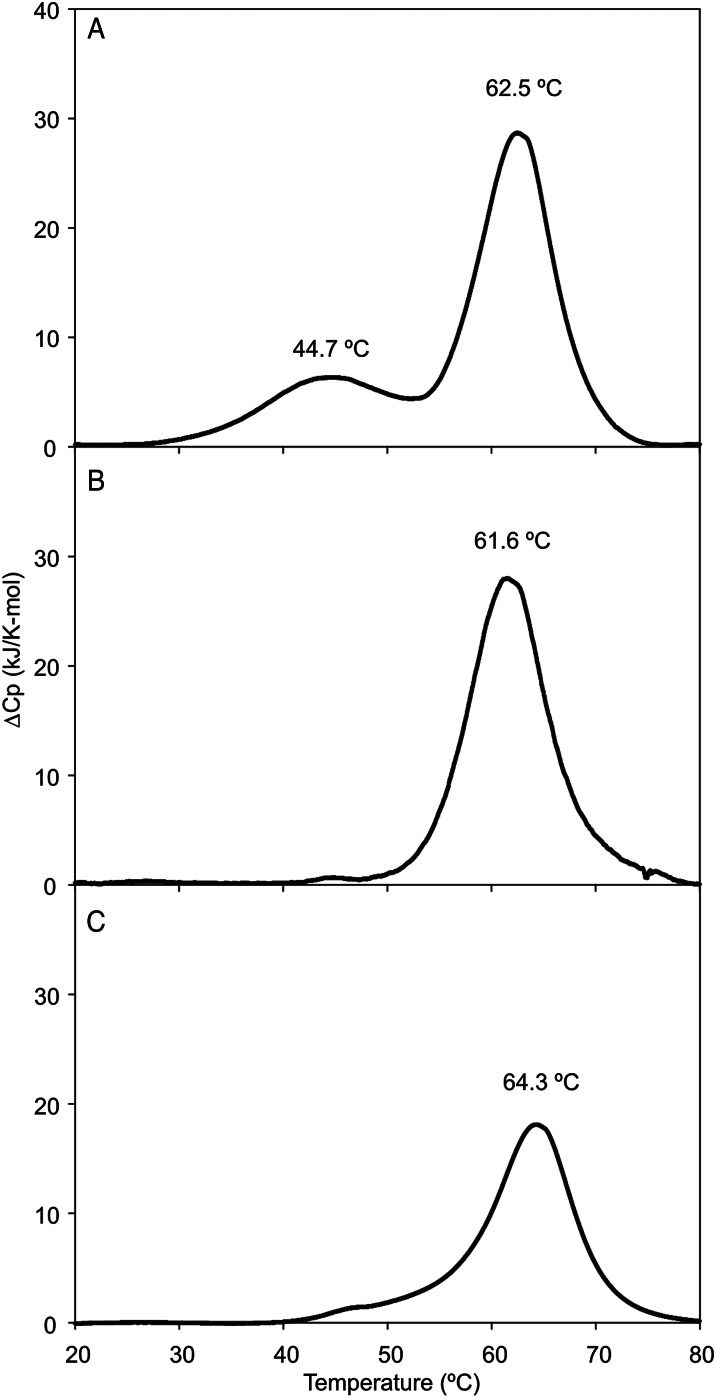
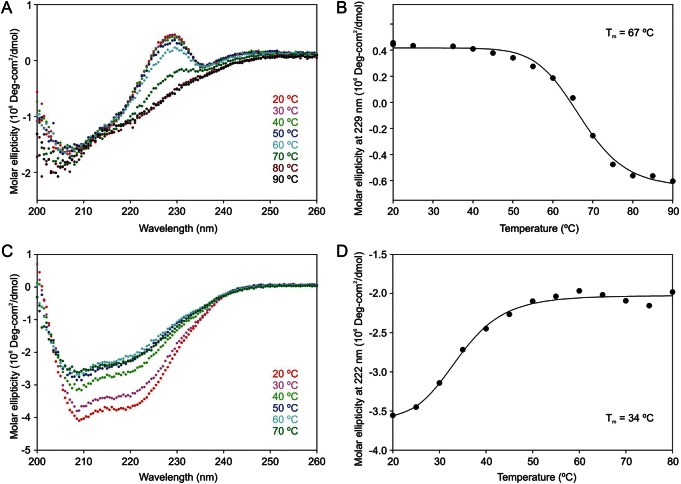
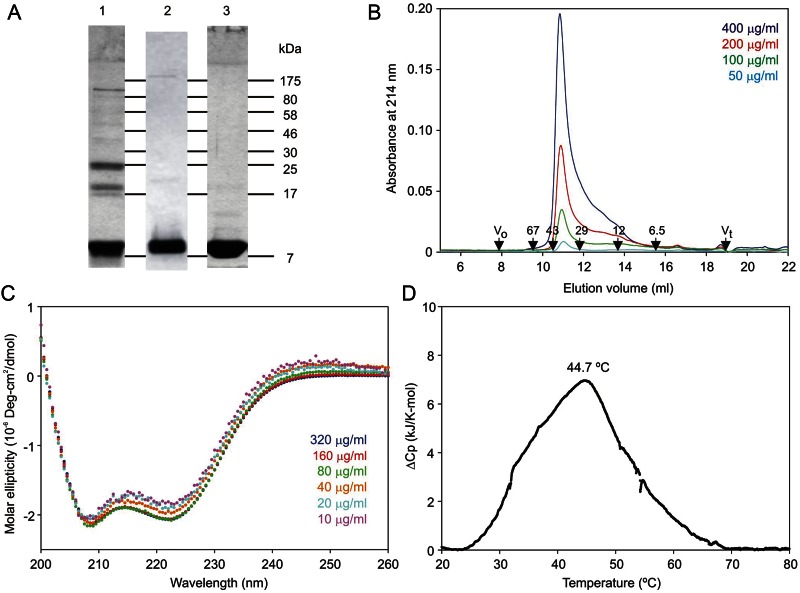
The properties of the human macrophage galactose receptor have been investigated. Specificity for N-acetylgalactosamine (GalNAc) residues with exposed 3- and 4-hydroxyl groups explains virtually all of the results obtained from a recently expanded array of synthetic glycans and is consistent with a model for the structure of the binding site. This simple interaction is sufficient to explain the ability of the receptor to bind to tumor-cell glycans bearing Tn and sialyl-Tn antigens, but not to more elaborate O-linked glycans that predominate on normal cells. This specificity also allows for binding of parasite glycans and screening of an array of bacterial outer membrane oligosaccharides confirms that the receptor binds to a subset of these structures with appropriately exposed GalNAc residues. A key feature of the receptor is the clustering of binding sites in the extracellular portion of the protein, which retains the trimeric structure observed in the cell membrane. Chemical crosslinking, gel filtration, circular dichroism analysis and differential scanning calorimetry demonstrate that this trimeric structure of the receptor is stabilized by an α-helical coiled coil that extends from the surface of the membrane to the globular carbohydrate-recognition domains. The helical neck domains form independent trimerization domains. Taken together, these results indicate that the macrophage galactose receptor shares many of the features of serum mannose-binding protein, in which clusters of monosaccharide-binding sites serve as detectors for a simple epitope that is not common on endogenous cell surface glycans but that is abundant on the surfaces of tumor cells and certain pathogens.
Keywords: Carbohydrate-recognition domain; Galactose receptor; Glycan array; Glycan-binding receptor; Lectin.
Figures
Similar articles
-
Crystal Structure of the Carbohydrate Recognition Domain of the Human Macrophage Galactose C-Type Lectin Bound to GalNAc and the Tumor-Associated Tn Antigen.Biochemistry. 2021 May 4;60(17):1327-1336. doi: 10.1021/acs.biochem.1c00009. Epub 2021 Mar 16. Biochemistry. 2021. PMID: 33724805
-
A Novel Mechanism for Binding of Galactose-terminated Glycans by the C-type Carbohydrate Recognition Domain in Blood Dendritic Cell Antigen 2.J Biol Chem. 2015 Jul 3;290(27):16759-71. doi: 10.1074/jbc.M115.660613. Epub 2015 May 20. J Biol Chem. 2015. PMID: 25995448 Free PMC article.
-
Molecular cloning and expression of cDNA encoding human macrophage C-type lectin. Its unique carbohydrate binding specificity for Tn antigen.J Immunol. 1996 Jan 1;156(1):128-35. J Immunol. 1996. PMID: 8598452
-
Tn antigen interactions of macrophage galactose-type lectin (MGL) in immune function and disease.Glycobiology. 2023 Dec 25;33(11):879-887. doi: 10.1093/glycob/cwad083. Glycobiology. 2023. PMID: 37847609 Free PMC article. Review.
-
Diverse glycosylation of MUC1 and MUC2: potential significance in tumor immunity.J Biochem. 1999 Dec;126(6):975-85. doi: 10.1093/oxfordjournals.jbchem.a022565. J Biochem. 1999. PMID: 10578046 Review.
Cited by
-
Defining the conformation of human mincle that interacts with mycobacterial trehalose dimycolate.Glycobiology. 2014 Dec;24(12):1291-300. doi: 10.1093/glycob/cwu072. Epub 2014 Jul 15. Glycobiology. 2014. PMID: 25028392 Free PMC article.
-
Immature O-glycans recognized by the macrophage glycoreceptor CLEC10A (MGL) are induced by 4-hydroxy-tamoxifen, oxidative stress and DNA-damage in breast cancer cells.Cell Commun Signal. 2019 Aug 27;17(1):107. doi: 10.1186/s12964-019-0420-9. Cell Commun Signal. 2019. PMID: 31455323 Free PMC article.
-
MUC1 Glycopeptide Vaccine Modified with a GalNAc Glycocluster Targets the Macrophage Galactose C-type Lectin on Dendritic Cells to Elicit an Improved Humoral Response.J Am Chem Soc. 2023 Jun 21;145(24):13027-13037. doi: 10.1021/jacs.2c12843. Epub 2023 Jun 6. J Am Chem Soc. 2023. PMID: 37279388 Free PMC article.
-
A Bitter Sweet Symphony: Immune Responses to Altered O-glycan Epitopes in Cancer.Biomolecules. 2016 May 3;6(2):26. doi: 10.3390/biom6020026. Biomolecules. 2016. PMID: 27153100 Free PMC article. Review.
-
CD301 and LSECtin glycan-binding receptors of innate immune cells serve as prognostic markers and potential predictors of immune response in breast cancer subtypes.Glycobiology. 2024 Apr 1;34(3):cwae003. doi: 10.1093/glycob/cwae003. Glycobiology. 2024. PMID: 38206856 Free PMC article.
References
-
- Blixt O, Head S, Mondala T, Scanlan C, Huflejt ME, Alvarez R, Bryan MC, Fazio F, Calarese D, Stevens J, et al. Printed covalent glycan array for ligand profiling of diverse glycan binding proteins. Proc Natl Acad Sci U S A. 2004;101:17033–17038. doi:10.1073/pnas.0407902101. - DOI - PMC - PubMed
-
- Burnette WN. Western blotting”: Electrophoretic transfer of proteins from sodium dodecyl sulfate-polyacrylamide gels to unmodified nitrocellulose and radiographic detection with antibody and radioiodinated protein A. Anal Biochem. 1981;112:195–203. doi:10.1016/0003-2697(81)90281-5. - DOI - PubMed
-
- Coombs PJ, Harrison R, Pemberton S, Quintero-Martinez A, Parry S, Haslam SM, Dell A, Taylor ME, Drickamer K. Identification of novel contributions to high-affinity glycoprotein–receptor interactions using engineered ligands. J Mol Biol. 2010;396:685–696. doi:10.1016/j.jmb.2009.11.073. - DOI - PMC - PubMed
-
- Coombs PJ, Taylor ME, Drickamer K. Two categories of mammalian galactose-binding receptors distinguished by glycan array profiling. Glycobiology. 2006;16:1C–7C. doi:10.1093/glycob/cwj126. - DOI - PMC - PubMed
-
- Dam TK, Brewer CF. Effects of clustered epitopes in multivalent ligand-receptor interactions. Biochemistry. 2008;47:8470–8476. doi:10.1021/bi801208b. - DOI - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
